G-квадруплексы
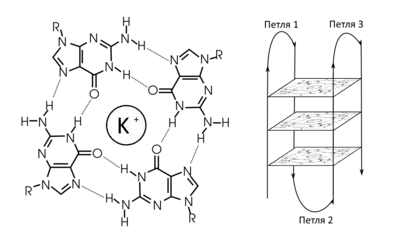
G-квадру́плексы (англ. G-quadruplex, а также G-tetrads или G4) — последовательности нуклеиновых кислот, обогащенные гуанином и способные образовывать структуры из четырёх цепей. Цепи нуклеиновых кислот из гуанозиновых олиго- и полинуклеотидов способны связываться друг с другом при наличии моновалентного катиона небольшого размера, чаще всего — калия. С помощью дифракционного анализа было показано, что такие поли(G)-нити представляют собой новый тип укладки ДНК, четырёхцепочечную спираль, где четыре гуаниновых основания из разных цепей образуют плоскую структуру, удерживаемую парными взаимодействиями G-G (рис. 1). Такие структуры отличаются высокой стабильностью в растворе и называются гуаниновыми (G)-квартетами, или G-тетрадами. Каждый G-квартет скреплен в сумме восемью водородными связями, образованными взаимодействием Уотсон-Криковской стороны одного гуанинового основания с Хугстиновской стороной другого. G-квадруплексы могут быть также образованы короткими олигонуклеотидами с соответствующей последовательностью, которую можно схематически записать как GmXnGmXoGmXpGm, где m — количество гуанинов в G-блоке. Эти гуанины обычно непосредственно задействованы в образовании G-тетрад. Xn, Xo и Xp могут быть комбинацией любых остатков, включая G; такие участки формируют петли между G-тетрадами.
Источники G-тетрадных мотивов
Нуклеиновые кислоты, содержащие G-тетрадный мотив, чрезвычайно широко представлены во всех открытых на данный момент геномах. Такие мотивы были обнаружены в промоторных регионах, интронах и сайтах переключения в составе последовательности генов иммуноглобулинов, «горячих точках» рекомбинации и др. При анализе генома человека было выявлено более 350 000 последовательностей, теоретически способных принимать конформацию квадруплексов. По-видимому, квадруплексы находятся в динамическом равновесии с другими формами ДНК, например, обычным дуплексом[1].
Теломерные квадруплексы
G-квартеты также представлены в ДНК на концах эукариотических хромосом, известных как теломеры. Теломерная ДНК представляет собой тандемные повторы коротких поли-G-блоков, которые иногда включают в себя адениловые или тимидиловые нуклеотиды: (GGTTAG)n, или (TTAGGG)n; при этом тип повтора является видозависимым: например, повтор (TTAGGG)n характерен для млекопитающих.
Функция теломер заключается в защите хромосомных концов от нежелательных повреждений в результате рекомбинации или воздействия нуклеаз. Человеческая теломерная ДНК в соматических клетках в среднем составляет 8-10 тысяч пар оснований. Терминальные же 100—200 нуклеотидов с 3'-конца представляют собой однотяжевой «хвост», конформационно ничем не ограниченный. В живых клетках этот «хвост» ассоциирован с белком POt1, в отсутствие же этого белка однотяжевая теломерная ДНК способна складываться и димеризоваться, формируя четырёхцепочечные шпильки, которые могут стабилизироваться формированием гуаниновых тетрад. Другой способ стабилизации такой ДНК — формирование внутримолекулярных G-квартетов путём многократного складывания.
Квадруплексы в промоторных регионах
Ряд участков ДНК в промоторных регионах генов человека способен принимать конформацию квадруплексов, тем самым обеспечивая регуляцию экспрессии генов. К белкам, гены которых могут регулироваться подобным образом, относятся, например, фактор транскрипции с-MYC[2], нарушение которого часто бывает связано с лимфомой Беркитта[3], протоонкогены RET[4], Bcl-2[5], c-Kit[6], фактор роста эндотелия сосудов[7] и др.
Квадруплексы в 5'-нетранслируемой области мРНК
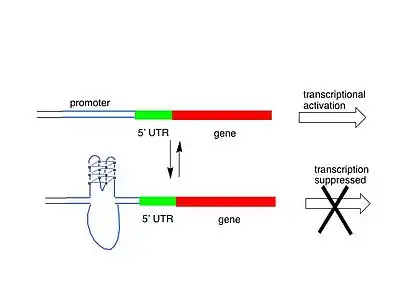
Биоинформатический анализ генома человека выявил, что около 3 000 мРНК содержат в своей 5'-нетранслируемой области один или несколько квадруплексов[9]. Квадруплекcы, расположенные в 5'-нетранслируемой области, могут участвовать в регуляции экспрессии генов на уровне трансляции[10]. Примерами таких матричных РНК человека являются мРНК рецептора эстрогена[11], внеклеточной металлопротеиназы[12], NRAS-протоонкогена[9] и др.
Синтетические квадруплексы
Нуклеиновые кислоты, как и белки, способны к избирательному связыванию различных молекул. Такие способные к специфическому связыванию олигонуклеотиды называют аптамерами. Относительно большой процент аптамеров содержит в своей структуре квадруплекс, который выполняет функцию стабилизации всей молекулы[13][14].
На сегодняшний день существует достаточно быстрый и эффективный способ получения ДНК- и РНК-аптамеров, способных связываться с практически любой более-менее крупной молекулой — SELEX. При помощи SELEX за последние 2 десятилетия было создано множество аптамеров, которые можно использовать для детекции различных веществ, а также в качестве основы для разработки лекарственных препаратов[15].
Примечания
- Huppert J. L., Balasubramanian S. Prevalence of quadruplexes in the human genome. (англ.) // Nucleic acids research. — 2005. — Vol. 33, no. 9. — P. 2908—2916. — doi:10.1093/nar/gki609. — PMID 15914667.
- Yang D., Hurley L. H. Structure of the biologically relevant G-quadruplex in the c-MYC promoter. (англ.) // Nucleosides, nucleotides & nucleic acids. — 2006. — Vol. 25, no. 8. — P. 951—968. — doi:10.1080/15257770600809913. — PMID 16901825.
- Spender L. C., Inman G. J. Developments in Burkitt's lymphoma: novel cooperations in oncogenic MYC signaling. (англ.) // Cancer management and research. — 2014. — Vol. 6. — P. 27—38. — doi:10.2147/CMAR.S37745. — PMID 24426788.
- Guo K., Pourpak A., Beetz-Rogers K., Gokhale V., Sun D., Hurley L. H. Formation of pseudosymmetrical G-quadruplex and i-motif structures in the proximal promoter region of the RET oncogene. (англ.) // Journal of the American Chemical Society. — 2007. — Vol. 129, no. 33. — P. 10220—10228. — doi:10.1021/ja072185g. — PMID 17672459.
- Agrawal P., Lin C., Mathad R. I., Carver M., Yang D. The major G-quadruplex formed in the human BCL-2 proximal promoter adopts a parallel structure with a 13-nt loop in K+ solution. (англ.) // Journal of the American Chemical Society. — 2014. — Vol. 136, no. 5. — P. 1750—1753. — doi:10.1021/ja4118945. — PMID 24450880.
- Hsu S. T., Varnai P., Bugaut A., Reszka A. P., Neidle S., Balasubramanian S. A G-rich sequence within the c-kit oncogene promoter forms a parallel G-quadruplex having asymmetric G-tetrad dynamics. (англ.) // Journal of the American Chemical Society. — 2009. — Vol. 131, no. 37. — P. 13399—13409. — doi:10.1021/ja904007p. — PMID 19705869.
- Sun D., Liu W. J., Guo K., Rusche J. J., Ebbinghaus S., Gokhale V., Hurley L. H. The proximal promoter region of the human vascular endothelial growth factor gene has a G-quadruplex structure that can be targeted by G-quadruplex-interactive agents. (англ.) // Molecular cancer therapeutics. — 2008. — Vol. 7, no. 4. — P. 880—889. — doi:10.1158/1535-7163.MCT-07-2119. — PMID 18413801.
- Bugaut A., Balasubramanian S. 5'-UTR RNA G-quadruplexes: translation regulation and targeting (англ.) // Nucleic Acids Res : journal. — 2012. — doi:10.1093/nar/gks068. — PMID 22351747.
- Kumari S., Bugaut A., Huppert J. L., Balasubramanian S. An RNA G-quadruplex in the 5' UTR of the NRAS proto-oncogene modulates translation. (англ.) // Nature chemical biology. — 2007. — Vol. 3, no. 4. — P. 218—221. — doi:10.1038/nchembio864. — PMID 17322877.
- Bugaut A., Balasubramanian S. 5'-UTR RNA G-quadruplexes: translation regulation and targeting. (англ.) // Nucleic acids research. — 2012. — Vol. 40, no. 11. — P. 4727—4741. — doi:10.1093/nar/gks068. — PMID 22351747.
- Balkwill G. D., Derecka K., Garner T. P., Hodgman C., Flint A. P., Searle M. S. Repression of translation of human estrogen receptor alpha by G-quadruplex formation. (англ.) // Biochemistry. — 2009. — Vol. 48, no. 48. — P. 11487—11495. — doi:10.1021/bi901420k. — PMID 19860473.
- Morris M. J., Basu S. An unusually stable G-quadruplex within the 5'-UTR of the MT3 matrix metalloproteinase mRNA represses translation in eukaryotic cells. (англ.) // Biochemistry. — 2009. — Vol. 48, no. 23. — P. 5313—5319. — doi:10.1021/bi900498z. — PMID 19397366.
- Tucker W. O., Shum K. T., Tanner J. A. G-quadruplex DNA aptamers and their ligands: structure, function and application. (англ.) // Current pharmaceutical design. — 2012. — Vol. 18, no. 14. — P. 2014—2026. — PMID 22376117.
- Gatto B., Palumbo M., Sissi C. Nucleic acid aptamers based on the G-quadruplex structure: therapeutic and diagnostic potential. (англ.) // Current medicinal chemistry. — 2009. — Vol. 16, no. 10. — P. 1248—1265. — PMID 19355883.
- Ni X., Castanares M., Mukherjee A., Lupold S. E. Nucleic acid aptamers: clinical applications and promising new horizons. (англ.) // Current medicinal chemistry. — 2011. — Vol. 18, no. 27. — P. 4206—4214. — PMID 21838685.
Литература
Книги
- Lucy W. Barrett, Sue Fletcher, Steve D. Wilton. Untranslated Gene Regions and Other Non-coding Elements. — SpringerBriefs in Biochemistry and Molecular Biology, 2013. — 57 p. — ISBN 978-3-0348-0679-4.
Статьи
- Решетников РВ, Копылов АМ, Головин АВ. Классификация G-квадруплексных ДНК по углу вращения квадруплекса и планарности G-квартетов // Acta Naturae : журнал. — 2010. — Т. 2, № 4. — С. 80—89. (недоступная ссылка)
- Guédin A., Gros J., Alberti P., Mergny J. How long is too long? Effects of loop size on G-quadruplex stability (англ.) // Nucleic Acids Research : journal. — 2010. — Vol. 32, no. 21. — P. 7858—7868. — doi:10.1093/nar/gkq639.
- Johnson J.E., Smith J.S., Kozak M.L., Johnson F.B. In vivo veritas: using yeast to probe the biological functions of G-quadruplexes (англ.) // Biochimie : journal. — 2008. — Vol. 90, no. 8. — P. 1250—1263. — doi:10.1016/j.biochi.2008.02.013. — PMID 18331848.
- Huppert JL and Balasubramanian S. Prevalence of quadruplexes in the human genome (неопр.) // NAR. — 2005. — Т. 33, № 9. — С. 2908—2916. — doi:10.1093/nar/gki609. — PMID 15914667.
- Todd A.K., Johnston M., Neidle S. Highly prevalent putative quadruplex sequence motifs in human DNA (неопр.) // NAR. — 2005. — Т. 33, № 9. — С. 2901—2907. — doi:10.1093/nar/gki553. — PMID 15914666.
- Burge S., Parkinson G.N., Hazel P., Todd A.K., Neidle S. Quadruplex DNA: sequence, topology and structure (неопр.) // NAR. — 2006. — Т. 34, № 19. — С. 5402—5415. — doi:10.1093/nar/gkl655. — PMID 17012276.
- Siddiqui-Jain A., Grand C.L., Bearss D.J., Hurley L.H. Direct evidence for a G-quadruplex in a promoter region and its targeting with a small molecule to repress c-MYC transcription (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2002. — Vol. 99, no. 18. — P. 11593—11598. — doi:10.1073/pnas.182256799. — PMID 12195017.
- Rawal P., Kummarasetti V.B., Ravindran J., Kumar N., Halder K., Sharma R., Mukerji M., Das S.K., Chowdhury S. Genome-wide prediction of G4 DNA as regulatory motifs: Role in Escherichia coli global regulation (англ.) // Genome Res. : journal. — 2006. — Vol. 16, no. 5. — P. 644—655. — doi:10.1101/gr.4508806. — PMID 16651665.
- Xu Hou, Wei Guo, Fan Xia, Fu-Qiang Nie, Hua Dong, Ye Tian, Liping Wen, Lin Wang, Liuxuan Cao, Yang Yang, Jianming Xue, Yanlin Song, Yugang Wang, Dongsheng Liu, and Lei Jiang. A biomimetic potassium responsive nanochannel: G-quadruplex DNA conformational switching in a synthetic nanopore (англ.) // J. Am. Chem. Soc. : journal. — 2009. — Vol. 131, no. 22. — P. 7800—7805. — doi:10.1021/ja901574c. — PMID 19435350.
- Quadruplex Nucleic Acids (неопр.) / Neidle & Balasubramanian. — 2006. — ISBN 0-85404-374-8. Архивная копия от 30 сентября 2007 на Wayback Machine