Тсиен, Роджер
Роджер Тсиен (Роджер Цянь, англ. Roger Tsien, кит. упр. 钱永健, пиньинь Qián Yǒngjiàn, палл. Цянь Юнцзянь[4]; 1 февраля 1952 — 24 августа 2016) — американский химик китайского происхождения, профессор кафедры химии и биохимии Калифорнийского Университета в Сан-Диего[5]. В 2008 году был удостоен Нобелевской премии по химии «за открытие и работу над зелёным флуоресцентным белком» совместно с двумя другими химиками[6].
| Роджер Тсиен | |
|---|---|
| англ. Roger Tsien | |
 | |
| Дата рождения | 1 февраля 1952[1][2][3][…] |
| Место рождения | |
| Дата смерти | 24 августа 2016[1][2][3][…] (64 года) |
| Место смерти | |
| Страна | |
| Научная сфера | биохимия |
| Место работы |
Калифорнийский университет в Беркли Калифорнийский университет в Сан-Диего |
| Альма-матер |
Гарвардский университет Кембриджский университет |
| Научный руководитель |
Richard Adrian Джереми Сандерс |
| Награды и премии |
|
| Сайт | tsienlab.ucsd.edu |
Биография
Ранние годы
Роджер Йончин Тсиен родился в Нью-Йорке 1 февраля 1952 года младшим из трех сыновей в семье Сюэчжу Тсиена, инженера-механика при Массачусетском Технологическом Институте, и медсестры Иин. Родословная его семьи связана с линией ученых-дворян из Ханчжоу, имеющей давние исторически документированные отношения с династией Сун. Более того, семья имела необычную судьбу, отражающую китайские исторические события во время и сразу после Второй мировой войны. Ее представители нередко делали карьеру на Западе, несмотря на неудачи и предрассудки. Так, Цянь Сюэсэнь, один из основателей Лаборатории реактивного движения при Калифорнийском технологическом институте, приходится двоюродным братом отцу Тсиена. Брат Тсиена, Ричард также является известным учёным в Стэнфорде. Роджер однажды сказал:
«Я от рождения обречён на такую работу».
Отец Роджера владел небольшой торговой компанией в Нью-Йорке, а также занимался инженерным консалтингом в округе Уэстчестер. Затем отец перевез семью в Ливингстон, штат Нью-Джерси, когда Роджеру исполнилось восемь лет. Там отец работал над вакуумными трубками и нефтехимикатами в Радиокорпорации Америки и Esso Research and Engineering.
В детстве Тсиен страдал от астмы и поэтому много времени проводил в помещениях. Он проявлял необычайно высокие способности к научному познанию, которые затем развились в любовь к теоретической и практической химии. Это ожидаемо началось с интереса к обусловленному лигандами поведению хромофоров — окрашенным переходам ионов металлов, участвующим в широком диапазоне классических экспериментов неорганической химии, и наиболее ярко проявило себя в попытках синтезировать ацетилсалициловую кислоту, часто на заднем дворе, в домашней посуде и с импровизированными приборами. Первый наградной значок бойскаута Роджер получил за заслуги в области химии. Его записная книжка, в которую он, восьмилетний ребенок, записывал свои химические опыты, теперь хранится в Музее Нобеля в Стокгольме[7].
Школьные годы
Талант к химии и любовь к ней сохранились у молодого Роджера и в школьные годы. Он прошел летнюю исследовательскую программу в Университете Огайо в 1967 году при поддержке Национального научного фонда. Исследование было посвящено амбидентным свойствам тиоцианата (SCN-), возникающим из схожего распределения отрицательного заряда между нуклеофильными атомами серы и азота. Это симметричное распределение заряда предполагало возможность образования связи, соединяющей два или более иона металла через связывание атома N и S c металлами класса А и В, соответственно. Несмотря на то, что Роджеру эта работа не принесла удовольствия и даже раздражала его, исследование, тем не менее, было отмечено высшей наградой в 1968 году в рамках конкурса «Поиск талантов в науке от компании Westinghouse», когда ему было 16 лет[7].
Развитие научной карьеры
Затем Роджер поступает в Гарвард, где его интерес к химии был прерван гуманитарным уклоном университета. В университете Тсиен встретился с Уолтером Гилбертом на курсах по молекулярной биологии, с Джеком Николсом, Дэвидом Хьюбелом (иностранный член Лондонского королевского общества, 1982) и Торстеном Визелем (иностранный член Лондонского королевского общества, 1982) на курсах по нейрофизиологии зрительных систем и с Нельсоном Киангом на лекциях по нейрофизиологии слуховых систем. Роджер вступил в общество Phi Beta Kappa по направлению химия и физика в 1972 году, закончив Гарвард summa cum laude (с отличием) с присуждением премии Детур (Detur Prize). Советы упомянутых выше преподавателей привели к тому, что Роджер успешно прошел конкурсный отбор на стипендию имени Маршала, покрывающую PhD-программу в Колледже Черчилля при Кембриджском университете, в физиологической лаборатории.
Руководитель его кембриджского PhD-проекта, профессор Ричард Эдриан интересовался клеточной физиологией активации скелетных мышц; они с Тсиеном стали друзьями. Эдриан был больше заинтересован в том, чтобы дать студентам высокого интеллектуального и научного калибра шанс развить свои оригинальные стремления и независимость, чем в наборе подчиненных, усердно и беспрекословно работающих под руководством главного исследователя. Так было в целом заведено в физиологической лаборатории, основанной Хиллом в 1965 году.
В научном подходе Тсиен унаследовал от Эдриана приверженность лабораторной работе и близкие взаимоотношения с небольшим числом коллег, которые обладали исключительными качествами, в небольшой исследовательской группе. Руководитель и студент поддерживали друг друга: спустя годы после окончания обучения, Роджер поддерживал контакт со своим наставником до конца жизни последнего.
Студента не привлекали имеющиеся электрофизиологические методы (методы регистрации особенностей внеклеточного ответа в отдельных нейронах внутри огромной популяции центральной нервной системы), применяющиеся в работе, хотя бы из-за их откликов на простые сенсорные стимулы. Он вёл долгие беседы с Ричардом Эдрианом по поводу применения преобразований Лапласа и кабельной теории дендритов в анализе электрического тока в клетках, способных к возбуждению; первое впоследствии легло в основу определений эффективной ёмкости в распределенной сети мембран[8], а обсуждения кабельной теории дали рождение новым методам «voltage clamp» в применении к бесконечным кабелям[9].
Роджер Тсиен близко общался с такими химиками, как Герри Смит, Йен Бэкстер и Джереми Сэндерс при Университетских химических лабораториях, а также с Арье Лью и Джоном Кимурой с коллегами из группы Дениса Хайдона при физиологической лаборатории. Таким образом, хотя Тсиен и не был экстравертом от природы, в университете он завёл много дружеских и профессиональных знакомств[7].
Последние годы и смерть
В 2014 году у Тсиена случился инсульт. Он переехал вместе со своей женой, Вэнди Глоуб, из Сан-Диего в Юджин, штат Орегон. Спустя два года, в 2016 году, в возрасте 64 лет Тсиен скончался во время езды на велосипеде.
Научное наследие Роджера Тсиена
Работы Тсиена были посвящены разработке различных методов измерения ключевых ионов и молекул в клеточной физиологии. Эти методы, в свою очередь, привели к созданию инструментов, которые позволили совершить важные явления в физиологии и расшифровать их механизмы уже при жизни учёного. Он также оставил после себя когорту выпускников и научных сотрудников[7].
Тсиен изучал широкий диапазон малых флуоресцентных молекул и флуоресцентных белков, которые привели к разработке важных методов прямой визуализации различных ключевых биохимических процессов внутри живой клетки и даже живых организмов. Эти вещества могут быть введены в клетку либо с помощью их способных к проникновению аналогов, либо путем их экспрессии напрямую в клетке с использованием молекулярно-биологических техник. Некоторые из них могут использоваться как индивидуальные флуоресцентные компоненты, некоторые — как взаимодействующие в паре (FRET). Это позволило применять их для изучения целого класса клеточных физиологических процессов, начиная от вопроса, включены ли гены, участвующие в этом процессе, переходя к изучению экспресии, транслокации и взаимодействия белков-участников, и заканчивая исследованием самих физиологических процессов, возникающих при взаимодействии белков[7].
Научные исследования
Аспирантура и дальнейшие исследования в Кембридже: биофизические методы слежения за физиологической активностью клетки
Диссертация Тсиена, завершенная в 1977 году, «Дизайн и использование химических инструментов в клеточной физиологии», была отмечена престижной премией Геджа в области биологии в Кембридже, а также предложением вступления в программу Comyns Berkeley Research Fellowship при Колледже Gonville and Caius. В этот период Тсиен начал разработки химических, а затем и молекулярно-биологических индикаторов, которые можно ввести в клетку или экспрессировать в ней для изучения её физиологических процессов. Данная задача сначала сводилась к измерению концентрации внутриклеточного кальция, хотя последующие работы включали уже гораздо более широкий диапазон ионов и молекул. Этот интерес Тсиена к измерению кальция проявился в наиболее подходящий момент времени: к концу 1970-х ученое сообщество постепенно признавало роль кальция в цитозоле как ключевого вторичного посредника. Прояснение его модификации, предшествующей триггерной активации или регуляторной стадии, и контроль этого процесса за счет обмена между свободными и связанными цитозольными компартментами, внутриклеточными кальциевыми депо и внеклеточным пространством, занимали центральное место в понимании механизма регуляции клеточной активности.
Разработка электрохимического метода обнаружения и количественного определения ионов кальция
Эта научная проблема возникла в процессе прямых измерений, проведённых с помощью электродов, способных проникать внутрь клетки, способной к возбуждению; такие измерения изначально применялись для определения мембранных потенциалов[10]. Электроды, селективные к ионам натрия, калия, водорода и хлорид-иону, к тому времени были уже доступны и нашли широкое применение[11], однако использование кальций-селективных электродов было ограничено рядом трудностей. К электродам предъявлялось требование высокой селективности по отношению к ионам кальция на фоне других катионов, потенциально находящихся в системе, в том числе катионов магния, натрия и водорода, а также требование высокой стабильности системы с минимальным гистерезисом при изменении концентрации кальция. Тсиен разработал более Ca2+-селективные, нейтральные (в противоположность к применяющимся анионам — органическим фосфатам) лиганды в качестве сенсоров[12]. Более того, требовались электроды с достаточно малым диаметром кончика (около 0,4 микрон)[13] с тем, чтобы избежать повреждения клеток, что могло привести к артефакту — повышению локальной концентрации кальция. Однако необходимые для этого высокие сопротивления, связанные с электродом, порядка 20-30 и 60-120 ГОм в растворе с р(Ca2+) = 3 и требующиеся для использования кончика с диаметрами 1,5 или 0,5 мкм, в свою очередь, повлекли за собой создание адаптированных, сделанных на заказ электрометров с очень высоким входным импедансом и малыми стабильными токами смещения (меньше 10 фА) во избежание ложных сигналов.
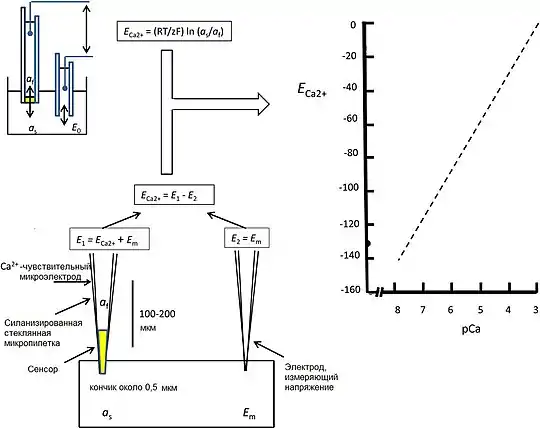
Экспериментальные данные, полученные с помощью оптимизированных электродов, удовлетворяли теоретически ожидаемому соотношению Нернста между выходным напряжением за вычетом одновременно записанных мембранных потенциалов и концентрацией свободного кальция вплоть до 1 мкМ в 0,1 М хлориде калия, с откликом, продолжающимся до 100 нМ [Ca2+] (рис. 1). Эти измерения согласовывались с данными, полученными путем уже устоявшейся люминесцентной спектроскопии в гигантских мышечных волокнах усоногих[14] и скелетных мышцах лягушки[15]. Измерения в последних были проведены с помощью флуорофора экворина, который содержит три Ca2+-связывающих EF-hand мотива. Экворин был получен из медузы Aequorea victoria, найденной в расположенной у западного побережья Северной Америки зоне Тихого Океана; вещество испускает флуоресцентную вспышку при возбуждении[16][17].
Оптические методы измерения Ca2+ с помощью разработанных органических молекул
Таким образом, электродные методы открыли дорогу количественному определению кальция в широких интервалах концентраций, обладали селективностью по отношению к кальцию на фоне ионов магния и водорода и являлись применимыми в электрофизиологических изысканиях. Однако для установления физиологической роли ионов кальция в клетке требовался метод, способный также к применению в широких диапазонах экспериментальных условий и типах клеток. В связи с этим было предложено несколько оптических подходов среди доступных флуоресцентных техник. Оптические методы показывают лучшую устойчивость сигнала к ионам натрия и более быстрый отклик по сравнению с электродными методами, что делало их подходящими для мониторинга клеточных событий. В отличие от измерений в конкретной точке, оптические методы также позволяли оценить концентрацию кальция, усредненную по какой-либо интересующей области, и охарактеризовать пространственное распределение ионов кальция при использовании доступной конфокальной микроскопии.
Исследования Тсиена при сотрудничестве с Тимоти Ринком и Тулио Поззаном внесли большой вклад в развитие оптических методов измерения и сделали их основным подходом, который применяется сегодня при изучении клеточной физиологии кальция. Было необходимо, чтобы разработанные для этой цели лиганды показали способные к обнаружению изменения в эмиссионных и абсорбционных свойствах в условиях возбуждения, совместимых с условиями записи сигнала и условиями жизнедеятельности клетки. Более того, молекулы должны обладать специфичностью к кальцию и предоставлять возможность для измерения как стационарных, так и промежуточных концентраций иона. Люминесцентный сигнал экворина обеспечивал воспроизводимые отклики на неравновесные концентрации, однако лиганд имел низкую аффинность к иону, а соотношение между концентрацией и сигналом выражалось уравнением со степенью 2,5, что сильно усложняло количественную интерпретацию эксперимента, особенно при изучении гетерогенных распределений иона в миоплазме[15]. Ставшие доступными позже флуорофоры обеспечивали более высокую аффинность, более широкий диапазон линейности и более быструю ассоциацию с ионами (антипирилазо-III, << 1 мс; дихлорофосфоназо-III, <2 мс; арсеназо-III, 2-3 мс) — это позволило отслеживать быстрые скачки кальция в скелетных мышцах[18]. Однако такие вещества, как арсеназо-III и антипирилазо-III показывали переменную стехиометрию связывания кальция, образуя комплексы 1:2 и 2:1 в зависимости от концентрации красителя; они также претерпевали большие изменения в своих адсорбционных свойствах в ответ на ионы магния и водорода в присутствии кальция[19]. Более того, наличие значительных концентраций индикатора могло само по себе повлиять на изучаемую систему: так, антипирилазо-III и арсеназо-III показали различающиеся временные зависимости поглощения после больших продолжительных импульсов напряжения около −20 мВ. Это дает основание полагать, что один или оба индикатора влияли на переходы кальция[20]. Вещество также потенциально могло связываться с цитоплазматическими компонентами или разделять их на нецитозольные компартменты — таковы возможные интерпретации, основанные на in vitro калибровках кюветы. Наконец, все эти индикаторы требовали введения в клетку, что напоминало микроэлектродные измерения. Это могло бы ограничить применение метода исследованием только достаточно сильных, хорошо закреплённых отдельных больших клетках, таких как мышечные волокна, гигантские аксоны кальмара и ретинальные клетки Limulus.
Возможность обеспечить специфическое связывание с кальцием в физиологическом диапазоне концентраций последнего возникла от идеи образования хелатного комплекса с четырьмя карбоксильными группами в хорошо известном лиганде этиленгликоль-бис(бета-аминоэтилэфир)-N,N,N`,N`-тетрауксусной кислоте (ЭГТА) (рис. 2а), образующем стехиометрический комплекс с ионом состава 1:1[21]. Это дало начало рациональному дизайну высокоаффинных буферов и оптических индикаторов кальция; первой ступенью в разработке стала замена метиленовых групп, связывающих кислород и азот, на фенильные остатки. В одном из таких аналогов, 1,2-бис(2-аминофенокси) этан-N,N,N`,N`-тетрауксусной кислоте (BAPTA), два бензольных кольца замещают метиленовые группы, соединяющие атомы N и O с сохранением суммарной геометрии, специфичности и аффинности молекулы к кальцию (рис. 2б). BAPTA — широко используемый и эффективный внутриклеточный кальциевый буфер, чье связывание по сравнению с ЭГТА меньше подвержено влиянию кислотности среды, более селективный по отношению к кальцию в среде магния и имеет большие скорости образования и разрушения комплекса.
Однако ЭГТА поглощает свет в дальней УФ-области и не флуоресцирует. BAPTA характеризуется УФ-спектром флуоресценции, который изменяется при связывании кальция, с максимумом около 250 нм, что не очень хорошо для флуоресцентного анализа. Было выяснено, что замена одного оксобензола на метоксихинолиновое кольцо привело в новом соединении хин-2 к появлению максимумов поглощения и испускания при 340 и 492 нм, соответственно (рис. 2с). Эти длины волн уже не менялись, однако интенсивность флуоресценции возрастала в шесть раз при связывании кальция. Измерения были откалиброваны при типичных концентрациях кальция в цитозоле в состоянии покоя (10−7 М) и ниже, вплоть до 10−8 М. Другие доступные методы того времени имели оптимум обнаружения только при концентрациях в активированном состоянии (10−6 М), тогда как сигналы в состоянии покоя были уже ниже предела обнаружения.
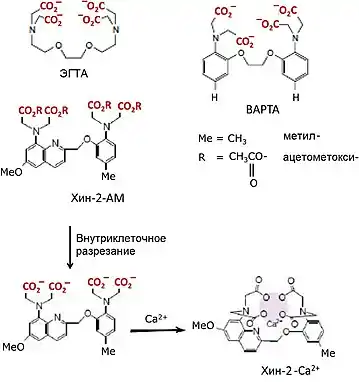
Наконец, инкубация клеток в растворах, содержащих проходящий через мембраны ацетометоксиэфирный фрагмент (АМ), связанный с хин-2 (хин-2-АМ), привела к атравматичному, постоянному доступу вещества внутрь клетки без микроманипуляций или нарушений мембраны. После попадания молекулы в клетку эндогенные эстеразы вырезали эфирный компонент и высвобождали активный, непроходящий через мембрану тетраанион (7) (рис 2г). Увеличение интенсивности флуоресценции с повышением концентрации кальция могло отслеживаться с применением обычного кюветного спектрофотометра. Таким образом, хин-2 приобрел ключевую роль в измерениях цитозольного кальция в широком диапазоне клеток млекопитающих и их суспензий, в том числе лимфоцитов, тромбоцитов, сперматозоидов, нейтрофилов и макрофагов, особенно при изучении роли кальция в процессе сигнал-ответ[22][23]. В будущем этот метод введения в клетку этерифицированных, проходящих через мембрану производных был расширен до изучения функций молекул — кандидатов на роль клеточных мессенджеров, таких как фосфатидилинозитол-3,4,5-трифосфат, в клеточной физиологии[24].
Калифорнийский университет в Беркли: новые лиганды, позволяющие измерять концентрацию внутриклеточного кальция и манипулировать им и другими ионами
Вероятно, в связи со сложностями в трудоустройстве и неясными перспективами работы, возникавших у ученых, занимавшихся междисциплинарными изысканиями в Великобритании, в 1981 году Тсиен после столь значительных исследований принял приглашение устроиться на работу в Отделение физиологии и анатомии в Калифорнийском университете в Беркли (в то время отделение возглавлял Терри Манчен) на ставку доцента. Там Тсиен проработал восемь лет, одним из направлений работы была дальнейшая разработка и оптимизация кальций-чувствительных лигандов в сотрудничестве со Стивеном Адамом и Робертом Зукером. Благодаря этому проекту в коммерческое производство были введены многие реагенты, которые сегодня крайне широко распространены в клеточной физиологии. Все они попадали в клетки после инкубации изучаемой системы с соответствующими АМ-производными. Вещества, соединявшие в себе 8-координационный тетракарбоксилат-хелатирующий фрагмент со стильбеновым хромофором, давали улучшенную селективность по отношению к кальцию на фоне других двухзарядных катионов и только немного меньшую аффинность, чем хин-2[21]. Аффинности этих веществ находились в диапазоне от Kd = 100 нМ (хин-2) до Kd = 90 мкМ (флуоро-5N), что покрывало типичные физиологические концентрации кальция в большом диапазоне типов возбужденных и покоящихся клеток. Более высокие значения длин волн возбуждения по сравнению с хин-2 (339 нм) позволяли избегать облучения клеток ультрафиолетом, который мог привести к автофлуоресценции возбужденной клетки и вызвать ее повреждения. Введение этиленовой группировки стильбена в гетероциклическое кольцо улучшило квантовый выход и фотохимическую стабильность — интенсивность флуоресценции могла увеличиться до 30 раз. Это улучшение в коэффициенте экстинкции и квантовом выходе по сравнению со значением в хин-2 (<5000 и 0,03 по сравнению с 0,14, соответственно) также уменьшило требуемое количество добавленной метки в систему (так, в случае хин-2 речь идет о миллимолярных концентрациях). Дело в том, что большие количества потенциально могли образовать буферную систему с внутриклеточным кальцием и нарушить изучаемый процесс.
Наконец, вдобавок к изменению интенсивности флуоресценции у веществ изменялись также и длины волн при связывании кальция, тогда как хин-2 давал изменения только в силе сигнала (это делало измерение чувствительным к вариациям в интенсивности облучения, детектировании сигнала, концентрации лиганда и эффективной толщине клетки в оптическом луче). Измерения с использованием спектральных сдвигов в области поглощения и/или испускания позволяли обходить эти источники артефактов[25]. Так, индо-1 имел двойной пик эмиссии с основным сдвигом от 475 до 400 нм при связывании кальция. Индо-1 нашел широкое применение в проточной цитометрии (рис. 3а). Фура-2 испускает только при одной длине волны 510 нм, но пик возбуждения сдвигается от приблизительно 380 до 350 нм при связывании, а отношение интенсивностей напрямую связано с концентрацией ионов. Высокий выход фотонов у этого соединения сделал его использование удобным даже при видеорегистрации локальной концентрации внутриклеточного кальция в режиме реального времени[26](рис. 3б).
Эти достижения дали начало созданию широкого класса новых молекул-детекторов разнообразных ионов и молекул, участвующих в физиологических процессах[27]. Во-первых, эти новые варианты позволили измерять концентрацию кальция в разных условиях и типах клеток. Сигнал флуо-3 регистрируется в видимой области при возбуждении аргоновым лазером при 488 нм, увеличиваясь при связывании при максимуме испускания на 525 нм, что близко к значениям, детектируемым в измерениях с флуоресцеин изотиоцианатом (FITC) (рис. 3в). Его более быстрая диссоциация по сравнению с фура-2 позволяет отслеживать быструю кинетику кальция в скелетных и сердечных мышцах. Это было особенно полезно в регистрации микроскопических событий высвобождения кальция ("кальциевые вспышки) при помощи конфокальной микроскопии[28]. Особенность также использовалась в регистрации результирующих скрытых распространяющихся Ca2+ волн, отражающих увеличенный выброс кальция рианодиновыми рецепторами в саркоплазматическом ретикулуме, который возникал из-за отсутствия сопряжения каналов с трансмембранным дигидропиридиновым рецептором в скелетных[29] или проаритмических обстоятельств в сердечных мышцах[30][31].
Во-вторых, стало возможно обнаружение и измерение концентрации не только для внутриклеточного кальция, но и для других участников процессов. Например, карбоксильные группы карбоксифлуоресцеинового производного 2`,7`-бис-(2-карбоксиэтил)-5-(и-6)-карбоксифлуоресцеин (BCECF) подходили скорее для определения ионов водорода, чем кальция. Доступность BCECF была достигнута за счет проникающего через клетки АМ-аналога, а само вещество имело один пик испускания (535 нм) и двойной пик возбуждения (около 490 и 440 нм). Его pKa (порядка 6.98) и линейный отклик между рН 6.4 и 7.4 обеспечили его применимость в клеточной физиологии в париетальных клетках желудка[32].
.jpg.webp)
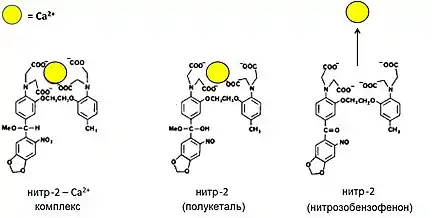
В-третьих, свет мог обратимо высвобождать кальций из Нитр-2 и позже разработанных нитр-5 и DM-нитрофена[33]. Нитр-2 состоит из кальций-хелатирующего BAPTA, соединенного с нитропиперонильной группой, которая способна претерпевать фотолиз светом из ближнего ультрафиолета (300—400 нм). Это событие значительно изменяет константу диссоциации кальция со 160 и 630 нМ до 7 и 18 мкМ при ионной силе в 0,1 и 0,3 М, соответственно, выпуская связанный кальций и изменяя, таким образом, концентрацию внутриклеточного кальция[34] (рис. 4а-в). Свойство нашло применение в фотохимическом управлении ионными токами в нейронах[35]. Затем Тсиен развил этот подход на другие биоактивные регуляторы, а пространственное разрешение было достигнуто фокусировкой лазерного источника возбуждения в трех координатах. Как результат, соединение бромированных 7-гидроксикумарин-4-илметилов с посредниками-кандидатами улучшило высвобождающий выход с применением двухфотонного, инфракрасного (в отличие от УФ) возбуждения, и позволило впервые получить трехмерную карту чувствительности к глутамату в срезах нейронов кортикального мозга крысы[36].
Разработка флуоресцентных белковых маркеров
_%D1%81_%D1%86%D0%90%D0%9C%D0%A4.jpg.webp)
Еще до переезда Тсиена в Сан-Диего в 1989, работа Александра Глейзера по флуоресцентным фикобилибелкам пробудила в нем интерес к флуоресцентному отслеживанию цАМФ. В качестве базы для флуоресцентной метки он рассматривал природные цАМФ-связывающие белки: это могло бы сразу обеспечить требуемые аффинности и селективности. В отсутствие цАМФ фосфокиназа А (ФКА) неактивна, а ее регуляторная и каталитическая субъединицы жестко связаны (рис. 5). Связывание цАМФ с регуляторными субъединицами приводит к диссоциации и активации каталитической, что дает возможность переноса фосфата из АТФ на некоторые определенные белки. Придание ферменту оптического сигнала в случае таких событий связывания возвратило Роджера к его университетским интересам в области флуоресцентного резонансного переноса энергии (FRET) между двумя светочувствительными хромофорами, находящимися стерически близко друг к другу. Возбужденный донорный хромофор может перенести энергию к акцептору за счет безызлучательного диполь-дипольного взаимодействия. Эта система была сконструирована за счет прикрепления флуорофора первого типа к регуляторной и второго типа к каталитической субъединицам. FRET мог возникнуть в интактной ФКА при близком контакте субъединиц двух типов, но должен был исчезнуть при диссоциации после связывания цАМФ: тогда сигналы в двух этих ситуациях наблюдались бы при разных длинах волн. Эти эксперименты, проведенные совместно с группой Сюзанны Тейлор, характеризовались тяжелой работой, требующей большой настойчивости: нужны были большие количества рекомбинантных субъединиц ФКА. Несмотря на трудности, проект закончился успехом и дал начало методу, в котором меченые флуоресцеином каталитические субъединицы связывались с мечеными родамином регуляторными субъединицами с образованием FRET-сенсоров на цАМФ[37]. Этот метод нашел применение в изучении остеобластов[38], меланоцитов[39] и нейронов Aplysia[40].
В университете Калифорнии в Сан-Диего: разработка генетически закодированных макромолекулярных индикаторов
Использование флуоресцентных белков, таким образом, элегантно дополнило класс низкомолекулярных лигандов. Однако необходимые белки нужно было в огромных количествах экспрессировать и очищать для селективного прикрепления двух разных меток in vitro к разным белковым доменам или субъединицам с одновременным сохранением функций белков. К тому же, в более ранних работах белки также надо было ввести в клетку. Эти трудности вынудили исследователей разрабатывать индикаторы, закодированные напрямую в геноме — в этом случае необходимо было только ввести гены, которые кодируют два флуоресцентных белка подходящего цвета, в изучаемые клетки. Такой подход предъявлял гораздо более низкие требования и подразумевал использование устоявшихся процедур для трансфекции клеток с малым количеством ДНК (по сравнению с введением белка) с последующим селективным отбором клеток, в которых прошла трансфекция. Эта задача обратила внимание Тсиена на медузу Aequorea victoria (уже второй раз), источник экворина[16], столь полезного в классических физиологических экспериментах. Шимомура обнаружил, выделил и очистил зеленый флуоресцентный белок (GFP) из приблизительно 10000 особей, с последующей характеризацией его физико-химических свойств, включающих спектральные, при разных условиях. Он показал, что GFP является FRET-акцептором возбуждения от донора-экворина и производит in vivo флуоресценцию в зеленой области спектра, отличную от синего сигнала очищенного препарата экворина при возбуждении в УФ-области[41]. Шимомура также определил хромофорную п-гидроксибензилиденимидазолиновую группировку в цепи белка[42].
Характеризация части гена GFP Дугласом Прэшером[43] инициировала совместную работу в разных лабораториях. Роджер работал над производством и флуоресцентными свойствами GFP, используя Saccharomyces cerevisiae в сотрудничестве с Роджером Хеймом и Скоттом Эмр. Мартином Чэлфи (иностранный член Лондонского королевского общества, 2018) из штата Колумбия, который первый продемонстрировал УФ-индуцированную флуоресценцию GFP, введённого в клетки беспозвоночного, и предложил потенциальное применение белка как биомаркера, работал над его экспрессией в Escherichia coli и Caenorhabditis elegan. Исследования Тсиена привели к обнаружению мутанта Y66H (BFP) с улучшенной стабильной флуоресценцией в синей области спектра по отношению к исходным GFP дикого типа. Дальнейшие модификации белка, стереохимически разместившие триптофан, привели к появлению мутанта Y66W, кодирующего циановый флуоресцентный белок (CFP)[44][45] (рис. 6а). В паре FRET конформационные изменения структуры УФ-возбужденного BFP приводили к переносу энергии к GFP (рис. 6б). Это подразумевало способность GFP возбуждаться волнами синего диапазона спектра, которые испускал донор BFP. Однако в спектре возбуждения GFP был большой пик в УФ- и маленький пик в синей области. Проблема была решена созданием мутантного варианта GFP: нежелательный пик в УФ-области исчез, а синий возрос приблизительно в 5-6 раз с последующим сдвигом +10 нм после мутации S65T[45]. В попытке проверить гипотезу исследователи ввели пептидную связь между BFP и мутантом GFP-S65T, а также с другими мутантами, имеющие схожий с GFP-S65T профиль возбуждения. Селективное УФ-возбуждение BFP, в самом деле, приводило к различным типам испускания в зеленой и синей области в присутствии или отсутствии FRET-переноса до и после трипсинолиза белка. Дальнейшие испытания подтвердили, что S65T является оптимальным флуорофором при условии введении мутации F64L, обеспечивающей сохранение структуры и фолдинг при более высоких температурах[46]. Этот двойной мутант, называемый «улучшенный (S65T-F64L) GFP», в больших объемах производится компанией Clontech.
Структурные исследования GFP-S65T выявили цилиндрическую структуру с диаметром 2,4 нм и длиной 4,0 нм, включающую 11 β-тяжей, окружающих вытянутую по оси спираль, в центре которой был вставлен хромофор[47] (рис. 7). Таким образом, группировка была защищена от воздействий растворителя и посторонних ферментов, но в пределах полости было пространство для потенциального введения ароматического кольца, которое связывалось бы за счет стекинга с хромофором; позже это было проверено посредством введения ряда мутаций, в том числе и T203Y, спектр этого варианта имел сдвиги возбуждения и испускания на приблизительно 20 нм. Получившийся желтый флуоресцентный белок (YFP) и его дальнейшие мутантные разновидности зарекомендовали себя как хорошие акцепторы сигнала от CFP, образуя альтернативные пары CFP/YFP.
_%D0%A1%D1%80%D0%B0%D0%B2%D0%BD%D0%B5%D0%BD%D0%B8%D0%B5_%D0%B0%D0%B1%D1%81%D0%BE%D1%80%D0%B1%D1%86%D0%B8%D0%BE%D0%BD%D0%BD%D1%8B%D1%85_%D0%B8_%D1%8D%D0%BC%D0%B8%D1%81%D1%81%D0%B8%D0%BE%D0%BD%D0%BD%D1%8B%D1%85_%D1%81%D0%B2%D0%BE%D0%B9%D1%81%D1%82%D0%B2_%D0%B2%D0%B0%D1%80%D0%B8%D0%B0%D0%BD%D1%82%D0%BE%D0%B2_GFP_%D0%B8_(%D0%B1)_%D0%B8%D1%85_%D1%81%D0%B1%D0%BE%D1%80%D0%BA%D0%B0_%D0%B2%D0%BE_FRET-%D0%BF%D0%B0%D1%80%D1%8B.jpg.webp)
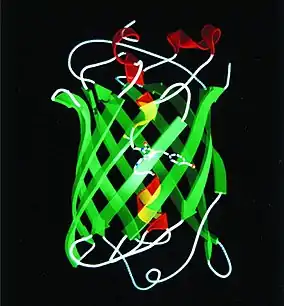
Роджер также подтвердил образование хромофорной п-гидроксибензилиденимидазолиновой группировки из остатков S65, Y66, и G67 в цепи GFP[31]. Механизм возникновения хромофора включал удивительное de novo образование гетероцикла с дегидрированием α-β С-С связи с образованием двойной связи. Для этого был необходим акцептор водорода — Тсиен полагал, что это был атмосферный кислород. Культивирование GFP-производящих бактерий в анаэробных условиях приводило к тому, что синтезированный белок не обладал флуоресценцией, однако она проявлялась несколько часов спустя после выдержки белка на воздухе[44] — свойство, которое затем нашло свое применение в изучении физиологии.
Создание белков-детекторов путем образования пар «донор FRET»-«акцепторный GFP-белок»
Применение FRET-визуализации специфических внутриклеточных сигналов требовало соединения компонентов FRET с молекулой, которая специфически связывала бы изучаемый клеточный компонент. В сотрудничестве с Ацуси Мияваки Роджер пытался прикрепить донорный и акцепторный белки к противоположным концам цитозольного домена недавно клонированного инозитол-1,4,5-трифосфатного (InsP3) рецептора. Скорее всего, эта работа являлась отражением его интереса в химическом сигналинге между клеточными мембранами. Проект привел к пониманию того, как этот важный посредник работает в скелетных мышцах в связке возбуждение-сокращение[48] и как работает фактор инфлюкса Ca2+[49], обеспечивающий трансдукцию из внутриклеточных депо Ca2+ к поверхности мембраны депо-оперируемых Ca2+ ворот[50]. В случае сигналинга, впоследствии было доказано, что процесс протекает под действием прямого, а не химического, сопряжения между поверхностными кальциевыми каналами L-типа и внутриклеточными саркоплазматическими ретикулярными рианодиновыми рецепторами выброса кальция[51]. Однако сложности, возникшие из-за неполного понимания механизма связывания InsP3 с рецепторами, заставили Тсиена обратить внимание на разработку других детекторов внутриклеточного кальция совместно с Мицусико Икура. Эта работа подразумевала прикрепление BFP и затем CFP к N-концу кальмодулина (CaM). S65T и затем YFP, наоборот, прикреплялись к С-концу целевого пептида М13. Конечная структура получалась соединением фрагментов СаМ и М13[52][53].
Получавшиеся таким образом генетически закодированные метки («хамелеоны»), способные к долгосрочному использованию и применимые к любой клетке или организму, в которые возможно введение таких ДНК, привели к появлению одного из самых популярных способов отслеживания активности в идентифицированных нейронах и расширили диапазон изучаемых объектов до интактных нервных систем. Более того, они раздвинули границы используемых биологически важных молекул. Значительные усилия исследователей привели к открытию линкеров, обеспечивающих слияние флуоресцентных белков с ФКА с одновременным сохранением способности субъединиц к ответу на присутствие цАМФ. Такие белки могли визуализировать субклеточное распределение цАМФ в кардиомиоцитах после стимуляции катехоламинами[54]. Наконец, модифицированный хамелеон, в котором М13 был заменен на пептид-субстрат киназы, а СаМ на белковый домен, содержащий фосфоаминокислотный связывающий домен, который может связывать фосфорилированный серин, треонин или тирозин, был способен визуализировать активность киназ, специфично фосфорилирующих серин, треонин или тирозин, соответственно. Фосфорилирование киназой указанных остатков приводит к образованию комплекса, в котором изменено расстояние или ориентация между донорным и акцепторным белками[55]. Вскоре этот метод стал незаменимым в исследовательской практике.
Завершение палитры цветов флуоресцентных белков; расширение области применения FRET для измерения мембранного потенциала
Дальнейшие разработки привели к пополнению арсенала флуоресцентных белков, покрыв широкий диапазон спектра и предоставив возможности к фотопереключению[56][57][58]. Открытие и доступность гена, кодирующего красный флуоресцентный белок из кораллов (DsRed)[59] заставили Роджера не ограничиваться GFP в своих исследованиях. DsRed является тетрамером, чья хромофорная составляющая начинается так же, как и в GFP. Однако дальнейшее дегидрирование приводит к образованию ацилимина, стабильного только в окружении интактного белка и показывающего длинноволновой сдвиг в спектрах поглощения и испускания[60][61]. Путем направленной эволюции был создан мономерный красный флуоресцентный белок (RFP), менее прихотливый к слиянию с другими белками по сравнению с тетрамером и ставший основой для целого набора мономерных белков, чьи максимумы испускания покрыли оставшуюся часть видимого спектра вплоть до 648 нм[62].
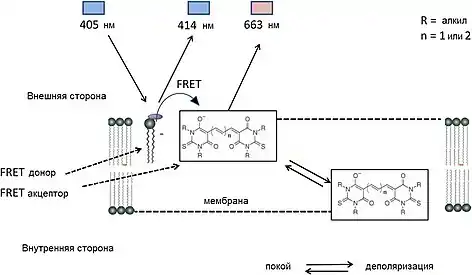
Роджер также приложил руку к манипуляциям с самой техникой FRET, реализовав свои ранние научные интересы в оптических измерениях мембранного потенциала. Это смогло улучшить устоявшиеся, но часто относительно медленные или низкочуствительные методы, основанные на одном индикаторном флуорофоре. Двухкомпонентный, основанный на FRET подход использовал в качестве донора флуоресцентные лектины, а позже меченый кумарином фосфатидилэтаноламин, связанный с внешней стороны мембраны (рис. 8). Доноры переносили энергию заряженному трансмембранному бис(1,3-дигексил-2-тиобарбитурат)три (или пента)метиноксонольному акцептору, когда он за счет заряда проходил из внешней стороны мембраны во внутреннюю при изменении мембранного потенциала. Этот подход позволял достичь беспрецедентной чувствительности (на каждые 100 мВ сигнал флуоресценции вырастал более чем на 50 %) и коротких временных постоянных (меньше 0,4 мс) по сравнению с другими оптическими индикаторами[63].
От клеточной физиологии к системной физиологии, трансляционной медицине и распространению новых индикаторов
Ближе к концу своей научной карьеры Тсиен сумел разработать такие инструменты оптического анализа, которые с особенным успехом нашли свое применение при исследовании целых органов или даже животных. Эти инструменты потенциально могли использоваться в областях клинической, нейрохирургической, сердечно-сосудистой или онкологической медицины. Некоторые из них были разработаны в сотрудничестве с Нгуен Тхао Куен (вьетн. Quyến Thẩm Nguyễn). Новая флуоресцентно-меченая легко вводимая в клетку и связывающаяся с нервом молекула F-NP41, направленная на взаимодействие с белком базальной мембраны ламинином, может улучшить оперативную визуализацию хронически усеченных нервов[64][65], что потенциально позволяет упростить разработку и хирургическое выполнение «ремонта» нервов[66][67]. Техника мечения для позитронно-эмиссионной томографии, использующая инъецированные эритроциты, меченые позитрон-испускающей, мультимодальной флуоресцентной меткой, может потенциально использоваться для обнаружения внутричерепного кровоизлияния как альтернатива 99mTc-меченым веществам[68] и может быть полезна в экспериментальных ЯМР-исследованиях церебральных патофизиологических процессов[69]. Введение в практику проникающих через мембрану пептидов, показывающих значительные изменения в флуоресценции после разрезания ассоциированными с опухолями протеазами внутри клетки может потенциально помочь в ранней диагностике злокачественных поражений[70]. Было выяснено, что как эти способные к активации пептиды, так и их связанные с гадолинием дендримерные формы селективно накапливаются в опухолевых клетках, что улучшает детекцию последних и предлагает средства для их диагностирования методом ЯМР в будущем[71]. Способные к активации пептиды похожим образом использовались для доставки анти-тубулинового радиосенсибилизатора монометилауристатина Е селективно к опухолевым клеткам, что открывает новый горизонт применения этих агентов в терапевтических целях[72].
На протяжении своей карьеры Тсиен пытался сделать разработанные им лиганды доступными для коллег. При ранних разработках хин-2 Роджеру не удалось убедить Национальную корпорацию исследований и разработок Соединенного королевства в коммерческой значимости обобщенной структуры, чувствительной к кальцию с беспрецедентной точностью и селективностью. Позже компания Lancaster Synthesis (ныне часть Johnson Matthey Company Alfa Aesar) заявила об интересе к хин-2 и его АМ-производному. Один из результатов научной активности Роджера — около 160 патентов в США. Тсиен также основал несколько компаний: так, с Чарльзом Зукером он создал Aurora Biosciences Corporation, производящую инструменты для поиска лекарств с помощью флуоресцентных маркеров, и Senomyx, занимающуюся поиском модуляторов вкусовых рецепторов. Благодаря его патентам другие люди тоже смогли образовывать компании, например, Molecular Probes.
Почести и награды
Нобелевская премия
Роджер разделил в 2008 году Нобелевскую премию в области химии с Осаму Симомурой (Морская биологическая лаборатория Вудс Хоул) и Мартином Чэлфи (Колумбийский университет, США). Формулировка комитета особенно подчеркнула вклад Тсиена в понимание флуоресцентных свойств GFP и близких к нему белков, который привел к их применению в исследовании динамических процессов в живых системах. В дань чести и уважения к Дугласу Прэшеру, который первым начал задачу изучения GFP, он пригласил Тсиена на церемонию.
Другие награды
- 1986 — Премия Лампорта Нью-Йоркской Академии Наук;
- 1989 — Награда Э. Йавица для исследователей в области нейронаук (NINCDS, 1989—1996);
- 1991 — Награда W. Alden Spencer
- 1995 — Международная награда Фонда Гайрднера;
- 1995 — Премия Artois-Baillet-Latour за достижения в области здравоохранения (Бельгия);
- 2002 — Награда Американского Химического Общества за креативные разработки;
- 2002 — Награда HP Heineken в области биохимии и биофизики, Королевская Нидерландская Академия Наук;
- 2002 — Медаль Макса Дельбрюка за достижения в молекулярной медицине, Институт Макса Дельбрюка, Берлин;
- 2004 — Премия Вольфа по медицине, Израиль;
- 2004 — Премия Кейо в области медицинских наук, Университет Кэйо, Япония;
- 2006 — Награда имени Льюиса С. Розенстила за выдающуюся работу в фундаментальной биомедицинской науке;
- 2006 — Медаль Уилсона;
- 2012 — Премия «Золотой гусь»
Членство в организациях и обществах
- 1995 — Избранный член Института медицины США;
- 1998 — Член Американской академии искусств и наук;
- 1998 — Член Национальной академии наук США;
- 2006 — Иностранный член Лондонского королевского общества;
- 2014 — Почетный член Кембриджского философского общества.
Семья
Тсиен был женат на Вэнди Глоуб. Родных детей у Роджера не было; с женой воспитывал пасынка Макса Ринка.
Личные качества
По словам его друга Кристофера Хуаня, Тсиен и его научный руководитель в аспирантуре Ричард Эдриан «разделяли схожие научные и личностные ценности, модели поведения, доброе, щедрое, уважительное отношение к миру и в высшей степени честное и человечное отношение к другим».
Увлечения и хобби
Занимался любительской фотографией. Любил активный отдых на природе.
См. также
Примечания
- https://www.washingtonpost.com/national/health-science/roger-y-tsien-chemist-shared-nobel-for-tool-to-research-alzheimers-dies-at-64/2016/08/31/90ca6de8-6fc9-11e6-8533-6b0b0ded0253_story.html
- Roger Y. Tsien // Encyclopædia Britannica (англ.)
- Roger Y. Tsien // Энциклопедия Брокгауз (нем.)
- 钱永健研水母发光盼助治癌. Lianhe Zaobao. Дата обращения: 8 октября 2008. (недоступная ссылка — история)
- Roger Y. Tsien at the University of California, San Diego (недоступная ссылка) (недоступная ссылка)
- Website of the Nobel Prize committee
- Huang C. L.-H. Roger Yonchien Tsien. 1 February 1952 — 24 August 2016 // Biogr. Mems Fell. R. Soc., 2018, v. 65, p. 405—428.
- Adrian R. H., Almers W. Measurement of membrane capacity in skeletal muscle.// Nat. New Biol., 1973, v. 242, p. 62-64.
- Adrian, R. H., Marshall, M. W. Sodium currents in mammalian muscle // J. Physiol.,1977, v. 268, p. 223—250.
- Adrian R. H. The effect of internal and external potassium concentration on the membrane potential of frog muscle. // J. Physiol., 1956, v. 133, p. 631—658.
- Thomas R. Ion-sensitive intracellular microelectrodes. New York, NY: Academic Press, 1978.
- Tsien R.Y., Rink T.J. Neutral carrier ion-selective microelectrodes for measurement of intracellular free calcium. // Biochim. Biophys. Acta Biomembr., 1980, v. 599, p. 623—638.
- Tsien R.Y. Liquid sensors for ion-selective microelectrodes. // Trends Neurosci., 1980, v. 3, p. 219—221.
- Tsien R.Y., Ashley C., Rink T. J. Changes in free Ca during muscle contraction, measured with an intracellular Ca-selective electrode // J. Physiol., 1978, v. 280, p. 27.
- Blinks J. R., Rüdel R., Taylor S. R. Calcium transients in isolated amphibian skeletal muscle fibres: detection with aequorin // J. Physiol., 1978, v. 277, p. 291—323.
- Shimomura O., Johnson F., Saiga Y. Extraction, purification and properties of aequorin, a bioluminescent protein from the luminous hydromedusan, Aequorea // J. Cell Comp. Physiol., 1962 v. 59, p. 223—239.
- Shimomura O. Luminescence of aequorin is triggered by the binding of two calcium ions // Biochem. Biophys. Res. Commun., 1995, v. 211, p. 359—363.
- Csernoch L., Huang C. L.-H., Szucs G., Kovacs L. Differential effects of tetracaine on charge movements and Ca2+ signals in frog skeletal muscle // J. Gen. Physiol., 1988, v. 92, p. 601—612.
- Baylor S. M. Optical studies of excitation-contraction coupling using voltage-sensitive and calcium-sensitive probes // Comp. Physiol., 2011 (оригинальная публикация: Handb. Physiol., Skelet. Muscle Suppl., 1983, v. 27, p. 355—379).
- Palade P., Vergara J. Arsenazo-III and antipyrylazo-III calcium transients in single skeletal muscle fibers // J. Gen. Physiol., 1982, v. 79, p. 679—707.
- Tsien R.Y. New calcium indicators and buffers with high selectivity against magnesium and protons: design, synthesis, and properties of prototype structures // Biochemistry, 1980, v. 19, p. 2396—2404.
- Tsien R.Y., Pozzan T., Rink T. J. T-cell mitogens cause early changes in cytoplasmic free Ca2+ and membrane potential in lymphocytes. Nature, 1982, v. 295, p. 68-71.
- Tsien R.Y., Pozzan T., Rink T. J. Calcium homeostasis in intact lymphocytes: cytoplasmic free calcium monitored with a intracellularly trapped fluorescent indicator // J. Cell Biol., 1982, v. 94, p. 325—334.
- Tsien R.Y., Jiang T., Sweeney G., Rudolf M. T. , Klip A., Traynor-Kaplan A. Membranepermeant esters of phosphatidylinositol 3,4,5-trisphosphate // J. Biol. Chem., 1998, v. 273, p. 11 017-11 024.
- Tsien R.Y., Grynkiewicz G., Poenie M. A new generation of Ca2+ indicators with greatly improved fluorescence properties // J. Biol. Chem., 1985, v. 260, p. 3440-3450.
- Cannell M. B., Berlin J. R., Lederer W. J. Intracellular calcium in cardiac myocytes: calcium transients measured using fluorescence imaging // Soc. Gen. Physiol. Ser., 1987, v. 42, p. 201—214.
- Haugland R. The molecular probes handbook: a guide to fluorescent probes and labeling technologies // . ThermoFisher Scientific, Waltham, MA USA, 2010 pp. 99-121. Режим доступа: https://www. thermofisher.com/uk/en/home/references/molecular-probes-the-handbook.html. Проверено 6 июня 2018.
- Cheng H., Lederer M. R., Xiao R. P., Gómez A. M., Zhou Y. Y., Ziman B., Spurgeon H., Lakatta E. G., Lederer W. J. Excitation-contraction coupling in heart: new insights from Ca2+ sparks // Cell Calcium, 1996, v. 20, p. 129—140.
- Chawla S., Skepper J. N., Hockaday A. R., Huang, C. L.-H. Calcium waves induced by hypertonic solutions in intact frog skeletal muscle fibres // J. Physiol., 2001, v. 536, p. 351—359.
- Goddard C. A., Ghais N. S., Zhang Y., Williams A. J., Colledge W. H., Grace A. A., Huang C. L.-H. Physiological consequences of the P2328S mutation in the ryanodine receptor (RyR2) gene in genetically modified murine hearts // Acta Physiol., 2008, v. 194, p. 123—140.
- Cody C. W., Prasher D. C., Westler W. M., Prendergast F. G., Ward W. W. Chemical structure of the hexapeptide chromophore of the Aequorea green-fluorescent protein // Biochemistry, 1993, v. 32, p. 1212—1218.
- Tsien R.Y., Paradiso A. M., Machen T. E. Na±H+ exchange in gastric glands as measured with a cytoplasmic-trapped, fluorescent pH indicator // Proc. Natl Acad. Sci. USA, 1984, v. 81, p. 7436-7440.
- Zucker R. S. Effects of photolabile calcium chelators on fluorescent calcium indicators // Cell Calcium, 1992, v. 13, p. 29-40.
- Tsien R.Y., Zucker R. S. Control of cytoplasmic calcium with photolabile tetracarboxylate 2-nitrobenzhydrol chelators // Biophys. J., 1986, v 50, p. 843—853.
- Gurney A. M.,Tsien R.Y., Lester H. A. Activation of a potassium current by rapid photochemically generated step increases of intracellular calcium in rat sympathetic neurons // Proc. Natl Acad. Sci. USA, 1987, v. 84, p. 3496-3500.
- Tsien R.Y., Furuta T. , Wang S. S.-H., Dantzker J. L., Dore T. M., Bybee W. J., Callaway E. M., Denk W. Brominated 7-hydroxycoumarin-4-ylmethyls: photolabile protecting groups with biologically useful cross-sections for two-photon photolysis // Proc. Natl Acad. Sci. USA, 1999, v. 96, p. 1193—1200.
- Tsien R.Y., Adams S. R., Harootunian A. T. , Buechler Y. J., Taylor S. S. Fluorescence ratio imaging of cyclic AMP in single cells // Nature, 1991, v. 349, p. 694—697.
- Tsien R.Y., Civitelli R. , Bacskai B. J., Mahaut-Smith M. P., Adams S. R., Avioli L. V. Single-cell analysis of cyclic AMP response to parathyroid hormone in osteoblastic cells // J. Bone Min. Res., 1994, v. 9, p. 1407—1417.
- Tsien R.Y., Sammak P. J. , Adams S. R. , Harootunian A. T., Schliwa M. Intracellular cyclic AMP, not calcium, determines the direction of vesicle movement in melanophores: direct measurement by fluorescence ratio imaging // J. Cell Biol., 1992, v. 117, p. 57-72.
- Tsien R.Y., Bacskai B. J., Hochner B., Mahaut-Smith M., Adams S. R., Kaang B. K., Kandel E. R. Spatially resolved dynamics of cAMP and protein kinase A subunits in Aplysia sensory neurons // Science, 1993, v. 260, p. 222—226.
- Morise H., Shimomura O., Johnson F. H., Winant J. Intermolecular energy transfer in the bioluminescent system of Aequorea // Biochemistry, 1974, v. 13, p. 2656—2662.
- Shimomura O. Structure of the chromophore of Aequorea green-fluorescent protein // FEBS Lett., 1979, v. 104, p. 220—222.
- Prasher D. C., Eckenrode V. K., Ward W. W., Prendergast F. G., Cormier M. J. Primary structure of the Aequorea victoria green-fluorescent protein // Gene, 1992, v. 111, p. 229—233.
- Heim T. R., Prasher D. C., Tsien R.Y. Wavelength mutations and posttranslational autoxidation of green-fluorescent protein. Proc. Natl Acad. Sci. USA, 1994, v. 91, p. 12501-12504.
- Tsien R.Y., Heim R. Engineering green-fluorescent protein for improved brightness, longer wavelengths and fluorescence resonance energy transfer // Curr. Biol., 1996, v. 6, p. 178—182.
- Cormack B. P., Valdivia R. H., Falkow, S. FACS-optimized mutants of the green-fluorescent protein (GFP) // Gene, 1996, v. 173, p. 33-38.
- Tsien R.Y., Brejc K., Sixma T.K. ,Kitts P.A. , KainS.R. , Ormo M., Remington S.J. Structural basis for dual excitation and photoisomerization of the Aequorea victoria green-fluorescent protein. Proc. Natl. Acad. Sci. USA, 1997, v. 94, p. 2306—2311.
- Tsien R.Y., Vergara J., Delay M. Inositol 1,4,5-trisphosphate: a possible chemical link in excitationcontraction coupling in muscle // Proc. Natl Acad. Sci. USA, 1985, v. 82, p. 6352-6356.
- Randriamampita C., Tsien R.Y. Degradation of a calcium influx factor (CIF) can be blocked by phosphatase inhibitors or chelation of Ca2+ // J. Biol. Chem., 1995, v. 270, p. 29-32.
- Putney J. W. Forms and functions of store-operated calcium entry mediators STIM and Orai // Adv. Biol. Regul., 2017, v. 68, p. 88-96.
- Huang C. L.-H., Pedersen T. H., Fraser J. A. Reciprocal dihydropyridine and ryanodine receptor interactions in skeletal muscle activation // J. Muscle Res. Cell Motil., 2011, v. 32, p. 171—202.
- Tsien R.Y., Miyawaki A., Llopis J., Heim R., Michael McCaffery J., Adams J. A., Ikura M. Fluorescent indicators for Ca2+ based on green-fluorescent proteins and calmodulin // Nature, 1997, v. 388, p. 882—887.
- Tsien R.Y., Miyawaki A., Griesbeck O.,Heim R. Dynamic and quantitative Ca2+ measurements using improved cameleons // Proc. Natl Acad. Sci. USA, 1999, v. 96, p. 2135—2140.
- Zaccolo M., Pozzan T. Discrete microdomains with high concentration of cAMP in stimulated rat neonatal cardiac myocytes // Science, 2002, v. 295, p. 1711—1715.
- Zhang J., Allen M. D. FRET-based biosensors for protein kinases: illuminating the kinome // Mol. Biosyst., 2007, v. 3, p. 759—765.
- Shaner N. C., Patterson G. H., Davidson M. W. Advances in fluorescent protein technology // J. Cell Sci., 2007, v. 120, p. 4247-4260.
- Tsien R.Y., Shaner N. C., Campbell R. E., Steinbach P. A. , Giepmans B. N. G., Palmer A. E. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein // Nat. Biotechnol., 2004, v. 22, p. 1567—1572.
- Tsien R.Y., Rodriguez E., Campbell R., Lin J. , Lin M. , Miyawaki A. , Palmer A. , Shu X., Zhang J. The growing and glowing toolbox of fluorescent and photoactive proteins // Trends Biochem. Sci., 2016 , v. 42, p. 111—129.
- Matz M. V., Fradkov A. F., Labas Y. A., Savitsky A. P., Zaraisky A. G., Markelov M. L., Lukyanov S. A. Fluorescent proteins from nonbioluminescent Anthozoa species // Nat. Biotechnol., 1999, v. 17, p. 969—973.
- Wall M. A., Socolich M., Ranganathan R. The structural basis for red fluorescence in the tetrameric GFP homolog DsRed // Nat. Struct. Biol., 2000, v. 7, p. 1133—1138.
- Yarbrough D., Wachter R. M., Kallio K., Matz M. V., Remington S. J. Refined crystal structure of DsRed, a red fluorescent protein from coral, at 2.0-Å resolution // Proc. Natl Acad. Sci. USA, 2001, v. 98, p. 462—467.
- Campbell R., Tsien R.Y., Tour. O., Palmer A., Steinbach P., Baird G. S., Zacharias D. A monomeric red fluorescent protein // Proc. Natl Acad. Sci. USA, 2002, v. 99, p. 7877-7882.
- Tsien R.Y., Gonzalez J. E. Improved indicators of cell membrane potential that use fluorescence resonance energy transfer // Chem. Biol., 1997, v. 4, p. 269—277.
- Hussain T., Tsien R.Y. et al. Fluorescently labeled peptide increases identification of degenerated facial nerve branches during surgery and improves functional outcome // PLoS ONE, 2015, v. 10, e0119600.
- Tsien R.Y. et al. Laminin targeting of a peripheral nerve-highlighting peptide enables degenerated nerve visualization // Proc. Natl. Acad. Sci. USA, 2016, v. 113, p. 12 774-12 779
- Glasby M. A., Gschmeissner S. G., Hitchcock R. J. I., Huang C. L.-H. The dependence of nerve regeneration through muscle grafts in the rat on the availability and orientation of basement membrane // J. eurocytol., 1986, v. 15, p. 497—510.
- Gattuso J. M., Davies A. H., Glasby M. A., Gschmeissner S. E., Huang C. L.-H. Peripheral nerve repair using muscle autografts: recovery of transmission in primates // J. Bone Jt Surg. Ser. B, 1988 v. 70, p. 524—529.
- Tsien R.Y. et al. 18F-positron-emitting/fluorescent labeled erythrocytes allow imaging of internal hemorrhage in a murine intracranial hemorrhage model // J. Cereb. Blood Flow Metab., 2017, v. 37, p. 776—786.
- Smith J. M., Bradley D. P., James M. F., Huang C. L.-H. Physiological studies of cortical spreading depression // Biol. Rev. Camb. Philos. Soc., 2006, v. 81, p. 457—481.
- Tsien R.Y. et al. Early detection of squamous cell carcinoma in carcinogen induced oral cancer rodent model by ratiometric activatable cell penetrating peptides // Oral Oncol., 2017, v. 71, p. 156—162.
- Tsien R.Y., Malone C. D. , Olson E. S. et al. Tumor detection at 3 tesla with an activatable cell-penetrating peptide dendrimer (ACPPD-Gd), a T1 magnetic resonance (MR) molecular imaging agent // PLoS ONE. (http://dx.doi.org/10.1371/journal.pone.0137104)
- Tsien R.Y. et al. Tumor radiosensitization by monomethyl auristatin E: mechanism of action and targeted delivery // Cancer Res., 2015, v. 75, p. 1376—1387.
Ссылки
- Фотографии Роджера на портале Wikimedia Commons (англ.)
- Веб-сайт лаборатории Тсиена в университете Сан-Диего
- Веб-страница Роджера на портале Кембриджского университета
| Словари и энциклопедии | ||||
|---|---|---|---|---|
| Генеалогия и некрополистика | ||||