Тирозинкиназа Брутона
Тирозинкиназа Брутона (англ. Bruton tyrosine kinase, BTK) — нерецепторная тирозинкиназа Tec-семейства протеинкиназ.
| Тирозинкиназа Брутона | |
|---|---|
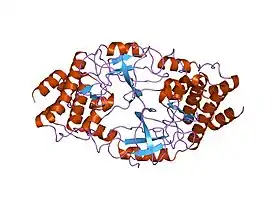 Структура каталитического домена тирозинкиназы Брутона | |
| Обозначения | |
| Символы | BTK; ATK, PSCTK1, XLA |
| Entrez Gene | 695 |
| HGNC | 1133 |
| OMIM | 300300 |
| PDB | 3GEN |
| RefSeq | NM_000061 |
| UniProt | Q06187 |
| Другие данные | |
| Шифр КФ | 2.7.10.2 |
| Локус | X-хр. , Xq21.33—22 |
Тирозинкиназа Брутона играет важную роль в созревании и функционировании В-лимфоцитов и других клеток крови млекопитающих. Наибольшее внимание уделяется изучению человеческого и мышиного вариантов фермента. Мутации в гене этого белка являются причиной болезни Брутона (X-сцепленной агаммаглобулинемии), откуда он и получил своё название. Тирозинкиназа Брутона является компонентом сигнальных путей, начинающихся с пре-B-клеточного и B-клеточного рецепторов. Учитывая большое значение этих путей для B-клеток, эту тирозинкиназу рассматривают как перспективную мишень при лечении хронического лимфолейкоза и аутоиммунных заболеваний[1].
История открытия и структура гена
Ген тирозинкиназы Брутона был идентифицирован в 1993 году двумя независимыми группами как ген, вовлечённый в развитие агаммаглобулинемии (болезни Брутона)[2][3]. Было показано, что ген локализован в длинном плече Х-хромосомы. Ген тирозинкиназы Брутона имеет размер около 37,5 тысяч пар нуклеотидов и содержит 19 экзонов[1]. Было идентифицировано более 1000 различных мутаций данного гена у пациентов с агаммаглобулинемией[4]. На матрице гена синтезируется мРНК размером 2,7 тысяч нуклеотидов. Данных о возможности альтернативного сплайсинга пре-мРНК тирозинкиназы в нормальных клетках пока нет. Однако при некоторых типах лейкоза обнаруживаются неправильно сплайсированные мРНК, на которых синтезируются молекулы тирозинкиназы Брутона, лишённые киназной активности[5].
Отсутствие функциональной формы тирозинкиназы у больных агаммаглобулинемией приводит к сильному снижению количества В-лимфоцитов и антител в крови. Аналогичные мутации в гене тирозинкиназы у мышей вызывают состояние X-сцепленного иммунодефицита. Его проявления менее выражены, чем у людей, например, количество периферических В-лимфоцитов снижено всего вдвое[6][7].
Распространение и внутриклеточная локализация
Тирозинкиназа Брутона характерна для В-лимфоцитов на разных стадиях их развития: в развивающихся клетках в костном мозге содержится больше всего белка, с выходом клеток в кровь его количество снижается, а в плазматических клетках фермент уже не обнаруживается[8][9][10]. Кроме того, тирозинкиназа Брутона присутствует и в других клетках костномозгового происхождения: моноцитах/макрофагах, тучных клетках, нейтрофилах, эритробластах, тромбоцитах, дендритных клетках и остеокластах[11][12][13][14][15][16][17][18]. Однако в Т-лимфоцитах эта киназа, по всей видимости, не синтезируется[10]. Тирозинкиназа Брутона играет более важную роль в судьбе В-лимфоцитов, чем любых других клеток. Это подтверждается тем, что мутации в гене фермента при болезни Брутона затрагивают в первую очередь созревание именно этих клеток[1]. В клетках других типов функции тирозинкиназы, по всей видимости, дублируются другими ферментами.
Белки, гомологичные тирозинкиназе Брутона млекопитающих, обнаружены также у птиц, рыб и насекомых[19][20][21]. Однако несмотря на эволюционную консервативность аминокислотной последовательности этих белков, их функции различаются в разных группах организмов. Например, у дрозофилы гомолог тирозинкиназы важен для развития гениталий самцов, в то время как у млекопитающих этот фермент участвует в развитии и функционировании клеток иммунной системы.
Тирозинкиназа Брутона локализуется, главным образом, в цитоплазме клетки. При активации фермент перемещается в плазматическую мембрану за счёт взаимодействия PH-домена (см. ниже) с её компонентами. Небольшое количество белка обнаруживается также в клеточном ядре, это позволяет предполагать, что тирозинкиназа циркулирует между ядром и цитоплазмой. Известно, что экспорт фермента их ядра обеспечивает экспортин Crm1[22].
Структура
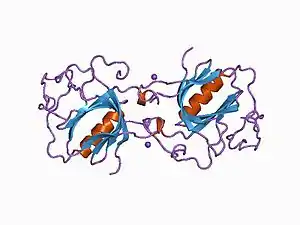
Тирозинкиназа Брутона человека состоит из 659 аминокислотных остатков и имеет молекулярную массу около 77 кДа. В белке выделяют следующие домены, начиная с N-конца: PH-домен (домен, гомологичный плекстрину), TH-домен (домен, гомологичный Tec; состоит из Btk-мотива и полипролинового региона), SH2-домен, SH3-домен и каталитический киназный домен[1].
PH-домен тирозинкиназы Брутона имеет характерную укладку и состоит из сильно изогнутого семитяжевого антипараллельного β-листа и С-концевой α-спирали. PH-домен связывает фосфатидилинозитолфосфаты цитоплазматической мембраны. Мутации в PH-домене, ассоциированные с болезнью Брутона, затрагивают аминокислотные остатки, непосредственно вовлечённые в связывание субстрата[23].
Btk-мотив соединён с С-концевой частью PH-домена линкером из 7—8 аминокислотных остатков. Глобулярное ядро Btk-мотива уложено напротив β-тяжей 5 и 7 PH-домена. В целом Btk-мотив представляет собой длинную петлю, которая складывается сама на себя и удерживается в таком положении за счёт иона цинка, формирующего координационные связи с остатками гистидина и цистеина. Ион цинка необходим для оптимальной активности и стабильности фермента. Одна из мутаций фермента, ассоциированных с болезнью Брутона, затрагивает один из цинксвязывающих остатков цистеина[23].
SH2-домен обеспечивает узнавание и связывание фосфотирозинсодержащих пептидов. SH3-домен связывает пролинбогатые участки других белков[24].
Регуляция
Количество и активность BTK регулируется на нескольких уровнях.
Регуляция на уровне транскрипции
Известно несколько факторов транскрипции, которые связываются и активируют промотор гена BTK: Sp1, Sp3, Spi-B, PU.1, OCT1/OBF1 и NF-κB (комплекс p65/p50). В последнем случае каталитическая активность BTK необходима для транслокации NF-κB в клеточное ядро, то есть имеет место авторегуляция тирозинкиназы Брутона по механизму положительной обратной связи[25].
Функции
Тирозинкиназа Брутона играет роль в развитии нескольких онкологических заболеваний, преимущественно лейкозов. Некоторые данные указывают на онкогенные свойства BTK, другие — на свойства супрессора опухолей[26].
Роль в развитии заболеваний
B-клеточный острый лимфобластный лейкоз
Инактивирующие соматические мутации гена BTK при B-клеточном остром лимфобластном лейкозе встречаются относительно редко[5][27]. Однако во многих случаях в лейкозных клетках обнаруживают аберрантно сплайсированные мРНК BTK, на которых синтезируются укороченные формы белка, лишённые киназного домена[28][29].
Ингибиторы
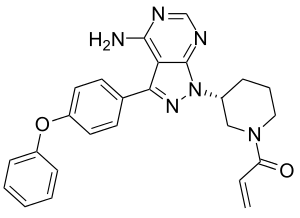
Понимание особой роли тирозинкиназы Брутона в патогенезе некоторых заболеваний привело к созданию ингибиторов её киназной активности с целью терапевтического применения. Одним из первых был разработан обратимый ингибитор киназ семейства Tec LFM-A13, который связывается с каталитической полостью тирозинкиназы, определяемой остатками Leu-460, Tyr-476, Arg-525 и Asp-539[30]. LFM-A13 ингибирует BTK в условиях in vitro c IC50=7,5 микромоль/литр. К 2013 году данных о клинической активности LFM-A13 получено не было[31].
К 2013 году были получены и другие обратимые ингибиторы BTK: ONO-WG-307, GDC-0834 и дазатиниб. ONO-WG-307 блокирует автофосфорилировние BTK по положению Tyr-223 с IC50=2 наномоль/литр. В доклинических испытаниях этот ингибитор показал эффективность в отношении диффузной B-крупноклеточной лимфомы подтипа ABC, фолликулярной лимфомы и хронического лимфолейкоза[31].
Дазатиниб был разработан как ингибитор тирозинкиназной активности BCR-ABL1 и применяется для лечения хронического миелобластного лейкоза и острого B-клеточного лимфобластного лейкоза, характеризующихся наличием химерного онкогена BCR-ABL1. Однако выяснилось, что он также обратимо ингибирует тирозинкинзу Брутона с IC50=5 наномоль/литр[32]. Дазатиниб был протестирован в клиническом исследовании II фазы на пациентах с рецидивирующим или устойчивым хроническим лимфолейкозом. В 20 % случаев был получен ответ на препарат, однако также были отмечены и частые серьёзные побочные эффекты[33].
Вторую группу ингибиторов BTK составляют ковалентные необратимые ингибиторы, такие как ибрутиниб (PCI-32765) и AVL-292 (CC-292). Обе молекулы образуют ковалентную связь с Cys-481 в активном сайте киназы. Эти ингибиторы характеризуются большей специфичностью в отношении тирозинкинзы Брутона и IC50=0,5 наномоль/литр.
Примечания
- Mohamed A.J., Yu L., Bäckesjö C.M., Vargas L., Faryal R., Aints A., Christensson B., Berglöf A., Vihinen M., Nore B.F., Smith C.I. Bruton's tyrosine kinase (Btk): function, regulation, and transformation with special emphasis on the PH domain (англ.) // Immunol Rev : journal. — 2009. — Vol. 228, no. 1. — P. 58—73. — doi:10.1111/j.1600-065X.2008.00741.x. — PMID 19290921.
- Tsukada S., Saffran D.C., Rawlings D.J., Parolini O., Allen R.C., Klisak I., Sparkes R.S., Kubagawa H., Mohandas T., Quan S., et al. Deficient expression of a B cell cytoplasmic tyrosine kinase in human X-linked agammaglobulinemia (англ.) // Cell : journal. — Cell Press, 1993. — Vol. 72, no. 2. — P. 279—290. — doi:10.1016/0092-8674(93)90667-F. — PMID 8425221.
- Vetrie D., Vorechovský I., Sideras P., Holland J., Davies A., Flinter F., Hammarström L., Kinnon C., Levinsky R., Bobrow M., et al. The gene involved in X-linked agammaglobulinaemia is a member of the src family of protein-tyrosine kinases (англ.) // Nature : journal. — 1993. — Vol. 361, no. 6409. — P. 226—233. — doi:10.1038/361226a0. — PMID 8380905.
- BTKbase Архивировано 16 февраля 2006 года.
- Feldhahn N., Río P., Soh B.N., Liedtke S., Sprangers M., Klein F., Wernet P., Jumaa H., Hofmann W.K., Hanenberg H., Rowley J.D., Müschen M. Deficiency of Bruton's tyrosine kinase in B cell precursor leukemia cells (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2005. — Vol. 102, no. 37. — P. 13266—13271. — doi:10.1073/pnas.0505196102. — PMID 16141323.
- Khan W.N., Alt F.W., Gerstein R.M., Malynn B.A., Larsson I., Rathbun G., Davidson L., Müller S., Kantor A.B., Herzenberg L.A., et al. Defective B cell development and function in Btk-deficient mice (англ.) // Immunity : journal. — Cell Press, 1995. — Vol. 3, no. 3. — P. 283—299. — PMID 7552994.
- Kerner J.D., Appleby M.W., Mohr R.N., Chien S., Rawlings D.J., Maliszewski C.R., Witte O.N., Perlmutter R.M. Impaired expansion of mouse B cell progenitors lacking Btk (англ.) // Immunity : journal. — Cell Press, 1995. — Vol. 3, no. 3. — P. 301—312. — PMID 7552995.
- Genevier H.C., Hinshelwood S., Gaspar H.B., Rigley K.P., Brown D., Saeland S., Rousset F., Levinsky R.J., Callard R.E., Kinnon C., et al. Expression of Bruton's tyrosine kinase protein within the B cell lineage (англ.) // European Journal of Immunology : journal. — 1994. — Vol. 24, no. 12. — P. 3100—3105. — doi:10.1002/eji.1830241228. — PMID 7805739.
- Nisitani S., Satterthwaite A.B., Akashi K., Weissman I.L., Witte O.N., Wahl M.I. Posttranscriptional regulation of Bruton's tyrosine kinase expression in antigen receptor-stimulated splenic B cells (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 2000. — Vol. 97, no. 6. — P. 2737—2742. — PMID 10688914.
- Smith C.I., Baskin B., Humire-Greiff P., Zhou J.N., Olsson P.G., Maniar H.S., Kjellén P., Lambris J.D., Christensson B., Hammarström L., et al. Expression of Bruton's agammaglobulinemia tyrosine kinase gene, BTK, is selectively down-regulated in T lymphocytes and plasma cells (англ.) // Journal of Immunology : journal. — 1994. — Vol. 152, no. 2. — P. 557—565. — PMID 8283037.
- Kawakami Y., Yao L., Miura T., Tsukada S., Witte O.N., Kawakami T. Tyrosine phosphorylation and activation of Bruton tyrosine kinase upon Fc epsilon RI cross-linking (англ.) // Molecular and Cellular Biology : journal. — 1994. — Vol. 14, no. 8. — P. 5108—5113. — doi:10.1128/MCB.14.8.5108. — PMID 7518558.
- Mukhopadhyay S., Mohanty M., Mangla A., George A., Bal V., Rath S., Ravindran B. Tyrosine phosphorylation and activation of Bruton tyrosine kinase upon Fc epsilon RI cross-linking (англ.) // Molecular and Cellular Biology : journal. — 1994. — Vol. 14, no. 8. — P. 5108—5113. — doi:10.1128/MCB.14.8.5108. — PMID 7518558.
- Futatani T., Miyawaki T., Tsukada S., Hashimoto S., Kunikata T., Arai S., Kurimoto M., Niida Y., Matsuoka H., Sakiyama Y., Iwata T., Tsuchiya S., Tatsuzawa O., Yoshizaki K., Kishimoto T. Deficient expression of Bruton's tyrosine kinase in monocytes from X-linked agammaglobulinemia as evaluated by a flow cytometric analysis and its clinical application to carrier detection (англ.) // Blood : journal. — American Society of Hematology, 1998. — Vol. 91, no. 2. — P. 595—602. — doi:10.1128/MCB.14.8.5108. — PMID 9427714.
- Quek L.S., Bolen J., Watson S.P. A role for Bruton's tyrosine kinase (Btk) in platelet activation by collagen (англ.) // Current Biology : journal. — Cell Press, 1998. — Vol. 8, no. 20. — P. 1137—1140. — doi:10.1016/S0960-9822(98)70471-3. — PMID 9778529.
- Robinson D., Chen H.C., Li D., Yustein J.T., He F., Lin W.C., Hayman M.J., Kung H.J. Tyrosine kinase expression profiles of chicken erythro-progenitor cells and oncogene-transformed erythroblasts (англ.) // J Biomed Sci. : journal. — 1998. — Vol. 5, no. 2. — P. 93—100. — PMID 9662068.
- Gilbert C., Levasseur S., Desaulniers P., Dusseault A.A., Thibault N., Bourgoin S.G., Naccache P.H. Chemotactic factor-induced recruitment and activation of Tec family kinases in human neutrophils. II. Effects of LFM-A13, a specific Btk inhibitor (англ.) // Journal of Immunology : journal. — 2003. — Vol. 170, no. 10. — P. 5235—5243. — PMID 12734372.
- Gagliardi M.C., Finocchi A., Orlandi P., Cursi L., Cancrini C., Moschese V., Miyawaki T., Rossi P. Bruton's tyrosine kinase defect in dendritic cells from X-linked agammaglobulinaemia patients does not influence their differentiation, maturation and antigen-presenting cell function (англ.) // Clinical and Experimental Immunology : journal. — 2003. — Vol. 133, no. 1. — P. 115—122. — doi:10.1046/j.1365-2249.2003.t01-1-02178.x. — PMID 12823285.
- Lee S.H., Kim T., Jeong D., Kim N., Choi Y. The Tec Family Tyrosine Kinase Btk Regulates RANKL-induced Osteoclast Maturation (англ.) // Journal of Biological Chemistry : journal. — 2008. — Vol. 283, no. 17. — P. 11526—11534. — doi:10.1074/jbc.M708935200. — PMID 18281276.
- Нуклеотидная последовательность мРНК BTK Gallus gallus
- Haire R.N., Strong S.J., Litman G.W. Tec-family non-receptor tyrosine kinase expressed in zebrafish kidney (англ.) // Immunogenetics : journal. — 1998. — Vol. 47, no. 4. — P. 336—337. — PMID 9472071.
- Baba K., Takeshita A., Majima K., Ueda R., Kondo S., Juni N., Yamamoto D. The Drosophila Bruton's tyrosine kinase (Btk) homolog is required for adult survival and male genital formation (англ.) // Molecular and Cellular Biology : journal. — 1999. — Vol. 19, no. 6. — P. 4405—4413. — PMID 10330180.
- Mohamed A.J., Vargas L., Nore B.F., Backesjo C.M., Christensson B., Smith C.I. Nucleocytoplasmic shuttling of Bruton's tyrosine kinase (англ.) // Journal of Biological Chemistry : journal. — 2000. — Vol. 275, no. 51. — P. 40614—40619. — doi:10.1074/jbc.M006952200. — PMID 11016936.
- Hyvönen M., Saraste M. Structure of the PH domain and Btk motif from Bruton's tyrosine kinase: molecular explanations for X-linked agammaglobulinaemia // EMBO J. — 1997. — Т. 16, вып. 12. — С. 3396—3404. — PMID 9218782.
- Uckun F. M. Bruton's tyrosine kinase (BTK) as a dual-function regulator of apoptosis // Biochem Pharmacol. — 1998. — Т. 56, вып. 6. — С. 683—691. — PMID 9751072.
- Yu L., Mohamed A. J., Simonson O. E., Vargas L., Blomberg K. E., Björkstrand B., Arteaga H. J., Nore B. F., Smith C. I. Proteasome-dependent autoregulation of Bruton tyrosine kinase (Btk) promoter via NF-kappaB (англ.) // Blood. — American Society of Hematology, 2008. — Vol. 111, iss. 9. — P. 4617—4626. — doi:10.1182/blood-2007-10-121137. — PMID 18292289.
- Hendriks R. W., Kersseboom R. Involvement of SLP-65 and Btk in tumor suppression and malignant transformation of pre-B cells // Semin Immunol. — 2006. — Т. 18, вып. 1. — С. 67—76. — PMID 16300960.
- Katz F. E., Lovering R. C., Bradley L. A., Rigley K. P., Brown D., Cotter F., Chessells J. M., Levinsky R. J., Kinnon C. Expression of the X-linked agammaglobulinemia gene, btk in B-cell acute lymphoblastic leukemia // Leukemia. — 1994. — Т. 8, вып. 4. — С. 574—577. — PMID 8152253.
- Goodman P. A., Wood C. M., Vassilev A. O., Mao C., Uckun F. M. Defective expression of Bruton's tyrosine kinase in acute lymphoblastic leukemia // Leuk Lymphoma. — 2003. — Т. 44, вып. 6. — С. 1011—1018. — PMID 12854903.
- Feldhahn N., Klein F., Mooster J. L., Hadweh P., Sprangers M., Wartenberg M., Bekhite M. M., Hofmann W. K., Herzog S., Jumaa H., Rowley J. D., Müschen M. Mimicry of a constitutively active pre-B cell receptor in acute lymphoblastic leukemia cells // J Exp Med. — 2005. — Т. 201, вып. 11. — С. 1837—1852. — PMID 15939795.
- Mahajan S., Ghosh S., Sudbeck E. A., Zheng Y., Downs S., Hupke M., Uckun F. M. Rational design and synthesis of a novel anti-leukemic agent targeting Bruton's tyrosine kinase (BTK), LFM-A13 [alpha-cyano-beta-hydroxy-beta-methyl-N-(2, 5-dibromophenyl)propenamide] // J Biol Chem. — 1999. — Т. 274, вып. 14. — С. 9587—9599. — PMID 10092645.
- Aalipour A., Advani R. H. Bruton tyrosine kinase inhibitors: a promising novel targeted treatment for B cell lymphomas // Br J Haematol. — 2013. — Т. 163, вып. 4. — С. 436—443. — doi:10.1111/bjh.12573. — PMID 24111579.
- Hantschel O., Rix U., Schmidt U., Bürckstümmer T., Kneidinger M., Schütze G., Colinge J., Bennett K. L., Ellmeier W., Valent P., Superti-Furga G. The Btk tyrosine kinase is a major target of the Bcr-Abl inhibitor dasatinib // Proc Natl Acad Sci U S A. — 2007. — Т. 104, вып. 33. — С. 13283—13288. — PMID 17684099.
- Amrein P. C., Attar E. C., Takvorian T., Hochberg E. P., Ballen K. K., Leahy K. .M., Fisher D. C., Lacasce A. S., Jacobsen E. D., Armand P., Hasserjian R. P., Werner L., Neuberg D., Brown J. R. Phase II study of dasatinib in relapsed or refractory chronic lymphocytic leukemia // Clin Cancer Res. — 2011. — Т. 17, вып. 9. — С. 2977—2986. — doi:10.1158/1078-0432.CCR-10-2879. — PMID 21402714.