Полони-секвенирование
Полони-секвенирование (англ. polymerase colony — «полимеразная колония») — технология секвенирования нового поколения (NGS) без электрофоретического разделения, позволяющая прочитывать миллионы коротких иммобилизованных последовательностей ДНК, значительно более дешевая по сравнению с секвенированием по Сэнгеру.
Технология была разработана в лаборатории доктора Джорджа Черча (George McDonald Church) в Гарварде (Гарвардская медицинская школа). По сравнению с другими методиками секвенирования — полони-секвенирование проводится на платформе с открытым исходным кодом и протоколами. Аппаратная платформа данного метода может быть легко комбинирована с широкодоступной эпифлуоресцентной микроскопией и микрофлюидными системами под управлением компьютера.
Основная идея метода — генерация большого числа уникальных 'полоний' (молекулярных колоний, сгенерированных полимеразой), которые секвенируются в случайном порядке. Полони-секвенирование проводится для библиотеки парных концевых тэгов (paired-end tags): каждая молекула ДНК имеет длину 135 п.о. (пар оснований), содержит два тэга длиной 17-18 п.о., разделённых и фланкированных общей последовательностью. Текущая длина прочтения для этой методики составляет 26 п.о. для ампликона и 13 п.о. для тэга (на каждом тэге остается непрочитанный зазор 4-5 п.о.).
Протокол
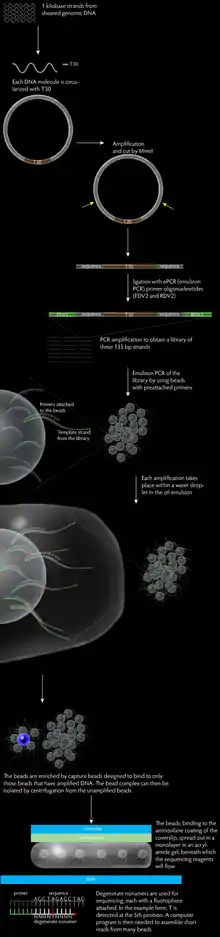
В протоколе полони-секвенирования можно выделить три основные стадии:
- конструкция библиотеки парных концевых тэгов
- амплификация
- секвенирование ДНК
Конструкция библиотеки парных концевых тэгов
Протокол начинается с фрагментирования исследуемой геномной ДНК на короткие последовательности (приблизительно равные по длине). Полученные короткие фрагменты подвергаются концевой репарации и концевой обработке аденином (добавление аденина к 3'-концу фрагментированной ДНК). Концевая репарация переводит любые поврежденные или выступающие концы ДНК в 5’-фосфорилированные тупые концы, тем самым обеспечивая немедленное лигирование по тупым концам. При помощи 6 % TBE ПААГ отбираются фрагменты ДНК длиной ~1000 п.о. На следующем этапе молекулы ДНК переводят в кольцевую форму с Т-концевыми синтетическими олигонуклеотидами длиной 30 п.о. (T30). T30 содержат два сайта для MmeI. Полученная кольцевая ДНК реплицируется по типу катящегося кольца. Амплифицированные кольцевые молекулы ДНК обрабатываются MmeI рестриктазой (эндонуклеаза рестрикции II типа), которая проводит разрезание на расстоянии от своего сайта узнавания. В результате рестрикции образуются T30 фрагменты, фланкированные с двух сторон тэгами длиной 17-18 п.о. каждый (общая длина фрагмента таким образом составляет ~70 п.о.). Полученные молекулы с парными концевым тэгами необходимо репарировать перед лигированием с праймерами (FDV2 и RDV2) к их концам для проведения эмульсионной ПЦР (еПЦР). Компоненты библиотеки — молекулы ДНК длиной 135 п.о. отбираются по размеру и подвергаются ник-трансляции. Заключительная стадия конструкции библиотеки — ПЦР-амплификация с целью увеличения количества материала библиотеки и элиминации посторонних продуктов реакции лигирования. Полученные матрицы ДНК включают последовательность FDV (44 п.о.), проксимальный тэг (17-18 п.о.), T30 (30 п.о.), дистальный тэг (17-18 п.о.) и последовательность RDV (25 п.о.).
Наиболее критический этап конструкции библиотеки концевых тэгов — получение кольцевых молекул геномной ДНК вокруг синтетического линкера. Эта стадия является скорость-лимитирующей для всего этапа.
Эмульсионная ПЦР
Парамагнитные шарики одинакового размера, покрыты стрептавидином и несут биотинилированные форвардные праймеры. Стрептавидин обладает очень сильной аффинностью к биотину, поэтому праймер очень прочно связан с поверхностью шарика. Водная фаза реакции содержит парамагнитные шарики, смесь компонентов для ПЦР, прямой и обратный праймеры, и библиотеку парных концевых тегов. Водная фаза перемешивается и взбалтывается с масляной фазой для получения эмульсии. В идеальной ситуации каждая капля воды в масляной эмульсии сдержит один шарик и одну молекулу матрицы ДНК, что позволяет в миллилитровом объёме проводить миллионы невзаимодействующих реакций ПЦР-амплификации.
Разбиение эмульсии
После амплификации, эмульсия, полученная на предыдущем этапе, разбивается обработкой изопропанолом и буферным раствором с детергентом (10 мМ Трис pH 7.5, 1мМ ЭДТА pH 8.0, 100 мМ NaCl, 1 % (об/об) Тритон X-100, 1 % (вес/об) SDS) и последующим встряхиванием, центрифугированием и магнитной сепарацией. Полученный раствор представляет собой суспензию с пустыми, клональными и неклональными шариками, которые получились из капель эмульсии без матрицы ДНК, с одной матрицей ДНК или с несколькими матрицами ДНК, соответственно. Обогащение шариками с амплифицированными продуктами проводят на следующей стадии.
Обогащение фракции шариками с ампликонами
Обогащение шариками с амплифицированными продуктами достигается за счёт гибридизации с более крупными, немагнитными шариками из полистирола, обладающих низкой плотностью. Полистироловые шарики несут биотинилированные олигонуклеотиды-ловушки (последовательности ДНК, комплементарные к eПЦР амплифицированным последовательностям). Смесь центрифугируют для отделения образовавшихся комплексов (микрошарики с ампликонами и шариками-ловушками) от микрошариков, не несущих ампликоны. Отделяемые комплексы имеют более низкую плотность и остаются в супернатанте, в то время как микрошарики без ампликонов образуют осадок. Супернатант отделяют и обрабатывают щёлочью (NaOH) для разрушения комплексов. Парамагнитные шарики с ампликонами отделяют от немагнитных шариков-ловушек при помощи магнитной сепарации. Такой протокол позволяет провести пятикратное обогащение шариками, несущими амплифицированные последовательности.
Кэпирование шариков
Присоединение кэпирущего олигонуклеотида к 3'-концам форвардных праймеров и RDV-сегменту матричной ДНК. Кэп содержит аминокислотную группу, которая препятствует лигированию флуоресцентного зонда к кэпированной последовательности, в то же время кэп способствует взаимодействию последовательности с аминосилированной поверхностью проточной ячейки.
Микрочип на покровном стекле
Покровные стекла моют и обрабатывают аминосиланом. Такая обработка способствует образованию ковалентных связей с матричной ДНК и устранению загрязнений флуоресцентными зондами. Фракцию микрошариков с ампликонами смешивают с акриламидом и выливают в небольшое углубление на предметном стекле, покрытое тефлоном. Предметное стекло с акриламидным гелем сразу же накрывают покровным стеклом, обработанным аминосиланом. Время полимеризации составляет 45 минут. Затем покровное и предметное стекло переворачивают и предметное стекло удаляют. Покровное стекло, обработанное силаном, ковалентно связывается с гелем, в то время как тефлон на поверхности предметного стекла обеспечивает лучшее отделение от акриламидного геля. Покровное стекло затем крепится к телу проточной ячейки и все несвязавшиеся микрошарики удаляются.
Секвенирование ДНК
Биохимическую основой полони-секвенирования служат дискриминационные свойства лигаз и полимераз. Сначала серии якорных праймеров прокачиваются через ячейку, праймеры гибридизуются с последовательностями синтетических олигонуклеотидов непосредственно на 3' или 5’-концах дистальных или проксимальных тэгах (длиной 17-18 п.о) геномной ДНК. Затем, проводится лигирование якорного праймера с вырожденными нонамерами (олигонуклеотиды длиной 9 н.т), меченными флуоресцентными метками.
5' Cy5‐NNNNNNNNT
5' Cy3‐NNNNNNNNA
5' TexasRed‐NNNNNNNNC
5' 6FAM‐NNNNNNNNG
Нонамеры, несущие флуорофоры, отжигаются с разным успехом к последовательностям тэгов в соответствии с методикой, напоминающей использование вырожденных праймеров. Однако, вместо презентации полимеразе, нонамеры избирательно лигируются на ДНК соседнего якорного праймера. Фиксация молекулы флуорофора обеспечивает сигнал флуоресценции, который позволяет различать присоединение A, C, G или T в исследуемой позиции геномного ДНК-тэга. После получения четырехцветных изображений комплексы между якорными праймерами нонамерными олигонуклеотидами смываются и начинается новый цикл за счёт замещения якорных праймеров. Вносится новая смесь флуоресцентно меченных нонамеров, для которых позиция, нуждающаяся в определении, сдвигается на 1 шаг по геномному тэгу.
5' Cy5‐NNNNNNNTN
5' Cy3‐NNNNNNNAN
5' TexasRed‐NNNNNNNCN
5' 6FAM‐NNNNNNNGN
Такая методика позволяет узнать последовательность 7 оснований в направлении от 5' к 3' концу и 6 оснований с 3'-конца. Окончательным результатом служит рид длиной 26 оснований (13 с каждого из концевых тэгов) с пропуском в 4-5 оснований в середине каждого тэга.
Временные затраты на выполнение протокола
Конструкция библиотеки концевых тегов занимает ~1 неделю, если длительные циклы инкубации проводить ночью. Протокол может быть остановлен по необходимости после любой стадии очистки продукта. Титрование концентрации матрицы для эмульсионной ПЦР занимает 2 дня. Эмульсионная ПЦР проводится за 2 дня. Запуск процесса секвенирования занимает 3 часа, каждый последующий цикл — 1.75 ч. Общее время непрерывных циклов секвенирования составляет 49 часов.
Анализ данных и программное обеспечение
В результате полони-секвенирования получаются миллионы чтений длиной 26 оснований за запуск. Эту информацию необходимо нормализовать и конвертировать в последовательности. Обработка осуществляется при помощи программного обеспечения, разработанного в лаборатории Черча (Church Lab). Всё программное обеспечение находится в открытом доступе.
Преимущества и недостатки метода
Полони-секвенирование позволяет получать последовательности ДНК с высокой точностью и высокой производительностью при помощи широкодоступного недорого инструмента. Кроме того, эта технология очень гибкая, что позволяет использовать её для различных целей, в том числе для секвенирования искусственных бактериальных хромосом, ресеквенирования бактериальных геномов а также для серийного анализа экспрессии генов (SAGE) и техник ДНК-штрихкода. Важным преимуществом полони-секвенирования является его небольшая стоимость. Технология полони-секвенирования может быть реализована при помощи широко доступного флуоресцентного микроскопа и поточной ячейки под управлением компьютера. Согласно расчетам на 2005 г., цена необходимого набора оборудования составляет приблизительно $130,000. Однако стоимость может быть уменьшена до $ 100,000 в ближайшем будущем. Технология полони-секвенирования реализована в приборе под названием полонатор. Стоимость прибора составляет $ 170,000. Согласно расчётам на 2005 г., цена каждой килобазы сырых данных составляет $0.11. Без учёта стоимости конструкции библиотеки парных концевых тэгов, цена чтения 1 килобазы может быть снижена до $0.08.
Несмотря на то, что получается большое число сырых первичных данных (786 гигабит), только один бит из 10,000 содержит полезную информацию. Другое затруднение — неравномерная амплификация снижает эффективность секвенирования и является на настоящий момент наибольшей сложностью для этой технологии.
Область применения
Полони-секвенирование наиболее эффективно при наличии референсного генома. Использовалось для ресеквенирования генома E. coli c точностью >99.9999 % и затратами в 9 раз меньше по сравнению с секвенированием по Сэнгеру. Технология полони-секвенирования применяется в платформе SOLiD (Applied Biosystems). PMAGE («polony multiplex analysis of gene expression» — множественный анализ экспрессии генов при помощи полони-технологии). Полони-секвенирование применяется для гаплотипирования и изучения сплайсинга.
Литература
1. Mitra, R. D., J. Shendure, et al. (2003). «Fluorescent in situ sequencing on polymerase colonies.» Anal Biochem 320(1): 55-65.
2. Shendure, J., G. J. Porreca, et al. (2005). «Accurate multiplex polony sequencing of an evolved bacterial genome.» Science 309(5741): 1728-32.
3. Porreca, G.J, Shendure, J., Church GM. (2006). «Polony DNA sequencing.» Curr Protoc Mol Biol: Chapter 7:Unit 7.8.
4. Lin B., Wang J., Cheng Y. (2008). «Recent Patents and Advances in the Next-Generation Sequencing Technologies.» Recent Pat Biomed Eng: (1):60-67.