Парабазалии
Парабаза́лии[1] (лат. Parabasalia) — группа гетеротрофных протистов из крупной клады Excavata, которой разные систематики присваивают ранг от класса до типа включительно. Характеризуются наличием особой структуры — парабазального аппарата, представленного аппаратом Гольджи, ассоциированным с поперечно исчерченными филаментами. Все представители, за редким исключением, имеют от 4 до десятков тысяч жгутиков. Помимо этого, для организмов данной группы характерно наличие в клетке мощного цитоскелетного образования — аксостиля, а также редукция митохондрий до специализированных органелл — гидрогеносом. Тип деления ядра — закрытый внеядерный плевромитоз.
| Парабазалии | ||||||||
|---|---|---|---|---|---|---|---|---|
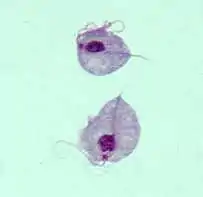 Trichomonas vaginalis | ||||||||
| Научная классификация | ||||||||
| Международное научное название | ||||||||
| Parabasalia Honigberg, 1973 | ||||||||
| Классы | ||||||||
|
||||||||
| ||||||||
Большинство парабазалий ведут симбиотический образ жизни, обитая внутри других организмов. Встречаются паразиты, в том числе возбудители заболеваний человека. Распространены повсеместно. По состоянию на 2012 год описано около 400 видов в 80 родах[2].
Биология

Все парабазалии, за редким исключением (некоторые виды из родов Pseudotrichomonas, Honigbergiella, Monotrichomonas и Ditrichomonas), ведут эндобиотический образ жизни, чаще всего обитая в отделах пищеварительного тракта животных[3]. Характер их взаимоотношений с хозяином может быть различным, от мутуалистических до паразитических. Широко известны симбиотические парабазалии (например, представители рода Trichonympha), обитающие в кишечнике термитов и родственных им тараканов рода Cryptocercus. Без симбионтов данные насекомые не способны усваивать целлюлозу. Виды родов Trichomonas, Dientamoeba, Histomonas являются широко распространёнными паразитами человека и других позвоночных. На останках тираннозавров были обнаружены повреждения, чрезвычайно сходные с наблюдаемыми у птиц при заражении Trichomonas gallinae, что свидетельствует о том, что парабазалии могли паразитировать на динозаврах[4].
Представители Parabasalia также часто образуют симбиотические ассоциации с бактериями. Последние могут располагаться как внутри клетки протиста и участвовать, например, в её метаболических процессах, так и прикрепляться снаружи. К примеру, на поверхности клетки Mixotricha paradoxa расположены сотни тысяч спирохет, выполняющих роль жгутиков, синхронное биение которых обеспечивает движение протиста, тогда как собственные 4 жгутика выполняют лишь рулевую функцию[5].
Строение
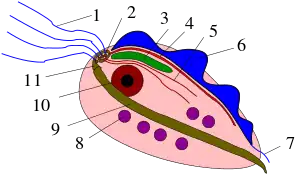
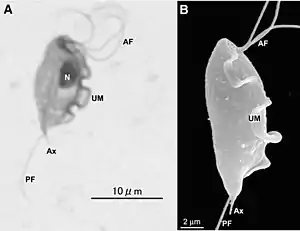
Строение, форма и размер клеток Parabasalia чрезвычайно варьируют в пределах таксона. Можно выделить два основных типа внешней организации клетки, характерные ранее выделяемым группировкам парабазалий: Trichomonadida и Hypermastigida. В первом случае клетка обычно имеет несколько жгутиков, отходящих от её переднего конца. Один жгутик направлен назад и формирует ундулирующую мембрану. Во втором случае клетка обычно несёт множество жгутиков, а ундулирующая мембрана отсутствует[6].
Ундулирующая мембрана, в случае её наличия, укреплена фибриллярным тяжем — костой. Эта структура встречается только у парабазалий. По ультратонкому строению фибрилл, входящих в состав косты, выделяют две основные разновидности данной органеллы — с исчерченностью A и B типа[3].
Для многих парабазалий характерно наличие особых электронно-плотных структур, ассоциированных с кинетосомами: инфра- и супракинетосомальных тел[3].
По середине клетки, от переднего к заднему концу, обычно располагается мощное микротрубочковое образование — пельта-аксостилярный комплекс. Пельта — органелла серповидной формы, поддерживающая кинетосомальный комплекс на переднем конце клетки. В зоне контакта с аксостилем в состав органеллы входят 35—40 микротрубочек. Аксостиль обычно представлен полым стержнем, проходящим по всей длине клетки и иногда выступающим за её пределы. Полость данной органеллы наполнена запасным веществом — гликогеном[7]. Пельта-аксостилярный комплекс может быть множественным, иногда слабо развит или отсутствует[3].
Уникальная клеточная структура парабазалий — парабазальный аппарат — состоит из совокупности парабазальных тел и отходящих от них парабазальных филаментов. Парабазальное тело представлено диктиосомами аппарата Гольджи. Обычно на одну диктиосому приходится 25—28 плотно прилегающих друг к другу цистерн, образующих замкнутые трубки, от которых отделяются пузырьки и гранулы с электронно-плотным содержимым. Парабазальный филамент всегда располагается параллельно стопке цистерн и состоит из 1—2 поперечно исчерченных фибрилл. Количество и расположение парабазальных тел с парабазальными филаментами в клетке может быть разным, часто коррелируя с организацией жгутикового аппарата[8].
В отличие от инфузорий, у парабазалий эволюционный процесс полимеризации жгутиков обычно не сопровождается увеличением количества ядер в клетке, хотя многоядерные формы также встречаются. Ядро, как правило, шаровидной формы, имеет одно, реже несколько ядрышек[3].
В цитоплазме парабазалий находятся многочисленные гидрогеносомы — сферические органеллы, покрытые двойной мембраной и заполненные гранулярным матриксом. Внутри данных органелл, по периферии располагаются от одной до нескольких кальций-содержащих везикул. Размер гидрогеносом в среднем составляет около 300 нанометров в диаметре, однако у Monocercomonas sp. может достигать 2 микрометров. В этих органеллах, являющихся редуцированными митохондриями, происходит анаэробное расщепление продуктов гликолиза с образованием АТФ и молекулярного водорода, сборка железосерных кластеров, там же локализуются некоторые компоненты системы синтеза аминокислот[9].
Геном
В 2007 году геном паразита человека Trichomonas vaginalis был расшифрован. Размер генома составил около 160 миллионов пар нуклеотидов, из которых около двух третей входят в состав повторяющихся последовательностей и транспозонов. По мнению авторов, подобная экспансия генома произошла при переходе данного вида от исходно анаэробного к микроаэрофильному местообитанию и, скорей всего, не характерна большинству парабазалий. В геноме T. vaginalis обнаружено около 60 тысяч белок-кодирующих последовательностей, при этом интроны найдены только в 65 из них. 152 гена предположительно были приобретены протистом в результате горизонтального переноса генов от бактерий. Большинство кодируемых ими белков вовлечены в метаболизм аминокислот и углеводов[10].
Особенности метаболизма
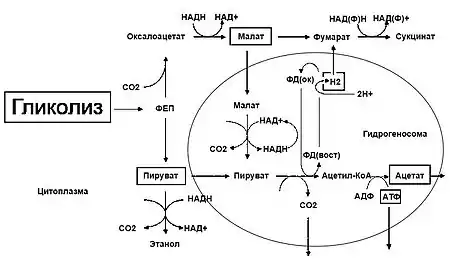
Система энергетического метаболизма парабазалий адаптирована к анаэробному или микроаэрофильному образу жизни, которые ведут эти протисты. Митохондрии представителей Parabasalia редуцировались до гидрогеносом, утративших собственный геном и большую часть компонентов электрон-транспортной цепи. За счёт данных органелл парабазалии способны продуцировать АТФ, разлагая продукты гликолиза, такие как малат и пируват, без участия кислорода. На первом этапе происходит окислительное декарбоксилирование пирувата, с образованием ацетил-КоА и углекислого газа, и переносом электрона на ферредоксин. Данная реакция катализируется ферментом пируват:ферредоксин-осидоредуктазой. Затем, за счёт работы гидрогеназы, электрон с ферредоксина переносится на протоны H+, с образованием молекулярного водорода. АТФ генерируется в результате двухстадийной реакции, с использованием энергии тиоэфирной связи, которая высвобождается при преобразовании ацетил-КоА в ацетат[9].
Жизненный цикл
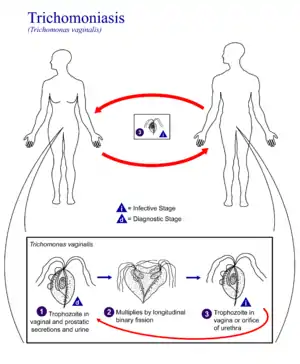
Жизненные циклы парабазалий чрезвычайно разнообразны. В качестве примера наиболее простого жизненного цикла можно привести таковой у паразита человека Trichomonas vaginalis, возбудителя венерического заболевания трихомониаз. Паразиты существуют только в виде пятижгутикового трофозоита, размножающегося бинарным делением. Передача от хозяина к хозяину происходит при половом контакте[11].
В жизненном цикле паразита птиц Histomonas meleagridis присутствуют амёбоидная и жгутиковая стадии. Первая поражает ткани печени и слепой кишки хозяев, тогда как жгутиковые формы обитают в просвете последней. Обе стадии способны к активному делению. Передача от хозяина к хозяину осуществляется за счет нематод рода Heterakis, также обитающих в слепой кишке птиц. Червь заглатывает гистомонад вместе с пищей, после чего те проникают в клетки кишечного эпителия нового хозяина и вновь приступают к активному питанию и размножению. Затем паразиты попадают в полость тела нематоды и из неё — в клетки репродуктивных органов, в результате чего яйца нематоды оказываются заражены Histomonas. В случае поедания вышедших вместе с фекалиями яиц другими птицами те заражаются как нематодой, так и протистом[12].
Жизненный цикл парабазалий — кишечных симбионтов таракана Cryptocercus (например, представителей родов Trichonympha и Urinympha) — включает в себя вегетативную фазу, которая осуществляется в нелиняющем хозяине и представляет собой длинную череду поколений, размножающихся бесполым путём, и половой процесс, который всегда происходит во время линьки хозяина. В ходе полового процесса всегда образуются цисты или псевдоцисты, способные сохранять жизнеспособность во внешней среде, куда большая часть симбионтов попадают вместе с отслоившейся кутикулой после линьки насекомого[13].
Характер полового процесса у разных видов сильно отличается. Вегетативная стадия видов рода Trichonympha гаплоидная. При подготовке хозяина к линьке данные организмы теряют большинство клеточных органелл и инцистируются. Под оболочкой цисты происходит формирование мужской и женской гамет. После линьки цисты, оставшиеся в кишечнике хозяина или поглощённые другими насекомыми, эксцистируются, происходит слияние гамет и образование зиготы. Последняя претерпевает двухступенчатый мейоз, что приводит к формированию 4 клеток вегетативного поколения. Такой тип полового процесса называется мерогамией[14].
В отличие от вышеописанного вида, у Urinympha вегетативная стадия диплоидна, и половой процесс протекает по типу автогамии. За 8 дней до начала линьки клетки данных организмов разбирают большинство клеточных органелл и преобразуются в псевдоцисты. На этой стадии проходит одноступенчатый мейоз с образованием двух дочерних ядер без деления самой клетки. Образовавшиеся ядра сливаются, и после окончания линьки хозяина псевдоцисты вновь преобразуются в вегетативные клетки[14].
Систематическое положение и классификация
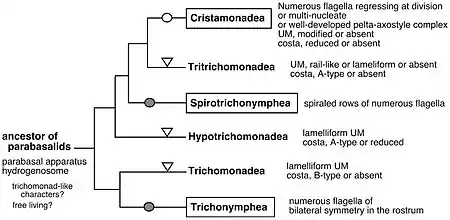
Таксон Parabasalia входит в группу эукариот Excavata, в подгруппу Metamonada[15]. Традиционно парабазалий рассматривали в качестве класса в типах Polymastigota или Tetramastigota, разделяя на два отряда: Trichomonadida (имеющих 4—6 жгутиков) и Hypermastigida (имеющих больше 10 жгутиков)[16][17]. В 2010 году Иван Чепичка (чеш. Ivan Čepička) с соавторами предложили рассматривать таксон в ранге типа. На основании молекулярно-филогенетических и морфологических данных авторы разделили парабазалий на 6 классов[3]:
- Hypotrichomonadea — наименьший по числу видов класс парабазалий. Представители характеризуются наличием 4 жгутиков, ундулирующей мембраны пластинчатовидного типа, двуветвистого парабазального тела, косты или основания косты с исчерченностью A-типа. Инфракинетосомальное тело отсутствует. Роды Hypotrichomonas, Trichomitus.
- Для Trichomonadea характерны 2—6 жгутиков, коста с исчерченностью B-типа, одиночный кариомастигонт (ядро и ассоциированные с ним кинетосомы), отсутствие инфракинетосомального тела. В этот класс, среди прочих, входят единственные представители парабазалий, ведущие свободноживущий образ жизни. Типичные представители: Trichomonas, Pentatrichomonas, Cochlosoma.
- Tritrichomonadea имеют от 0 до 5 жгутиков, супра- и инфракинетосомальным телом (за исключением представителей семейства Dientamoebidae). Коста, если присутствует, с исчерченностью A-типа. Могут быть одноядерными или двухъядерными. Типичные представители: Tritrichomonas, Dientamoeba, Histomonas.
- У представителей Cristamonadea может быть от одного до множества ядер. Кариомастигонт и аксостиль бывают одиночными, множественными или отсутствуют. Жгутиков от 2 до нескольких тысяч. Типичные представители: Lophomonas, Joenia, Devescovina.
- Виды, принадлежащие к классу Trichonymphea, разделены на передний рострум и задний пост-ростральный отдел. Жгутики обычно располагаются на особых жгутиковых зонах на роструме по несколько тысяч штук. Множество маленьких аксостилей не выступают за пределы клетки. Ядро одиночное. Типичные представители: Hoplonympha, Trichonympha.
- Spirotrichonymphea по внутреннему строению похожи на Trichonymphea, однако множество жгутиков располагаются на их клетках по спирали, закрученной против часовой стрелки. Типичные представители: Spirotrichonympha, Holomastigotoides.
В системе живых организмов, предложенных в 2015 году Руджеро с соавторами, таксон Parabasalia не выделяется, а всё разнообразие данной группы рассматривается в рамках классов Trichonymphea и Trichomonadea, входящих в тип Metamonada[18].
Медицинское значение
Среди парабазалий встречаются возбудители заболеваний человека, домашних и сельскохозяйственных животных. Tritrichomonas foetus поражает мочеполовую систему быков, а также вызывает хроническую диарею у домашних кошек[19]. Histomonas meleagridis паразитирует в пищеварительной системе и печени птиц, вызывая тяжёлые, часто смертельные заболевания у кур и индеек[20]. ''Dientamoeba fragilis'' может являться причиной болей в животе и диареи у человека[21]. Зарегистрированы случаи тяжёлых лёгочных заболеваний, вызываемых Trichomonas tenax[22].
Самым распространенным паразитом человека среди парабазалий является вид Trichomonas vaginalis (влагалищная трихомонада), вызывающий венерическую болезнь — урогенитальный трихомониаз. По данным ВОЗ, ежегодно в мире регистрируются более 174 миллионов случаев заболевания этой инфекцией[23].
Примечания
- Суханова, Бобылева, 2000, с. 357.
- Noda S, Mantini C, Meloni D, Inoue J, Kitade O, Viscogliosi E, Ohkuma M. Molecular phylogeny and evolution of parabasalia with improved taxon sampling and new protein markers of actin and elongation factor-1α // PLoS One. — 2012. — Vol. 7. — P. e29938. — doi:10.1371/journal.pone.0029938.
- Cepicka I, Hampl V, Kulda J. Critical taxonomic revision of Parabasalids with description of one new genus and three new species // Protist. — 2010. — Vol. 161. — P. 400—433. — doi:10.1016/j.protis.2009.11.005.
- Wolff EDS, Salisbury SW, Horner JR, Varricchi DJ. Common Avian Infection Plagued the Tyrant Dinosaurs // PLoS ONE. — 2009. — Vol. 4. — P. e7288. — doi:10.1371/journal.pone.0007288.
- Хаусман и др., 2010, с. 73.
- Хаусман и др., 2010, с. 71—74.
- Суханова, Бобылева, 2000, с. 362.
- Суханова, Бобылева, 2000, с. 377—378.
- Benchimol M. Hydrogenosomes under microscopy // Tissue and Cell. — 2009. — Vol. 41. — P. 151—168. — doi:10.1016/j.tice.2009.01.001.
- Carlton M., Hirt R., Silva J., Delcher A. et al. Draft Genome Sequence of the Sexually Transmitted Pathogen Trichomonas vaginalis // Science. — 2007. — Vol. 315. — P. 207—212. — doi:10.1126/science.1132894.
- Trichomoniasis. Centers for Disease Control and Prevention.
- Суханова, Бобылева, 2000, с. 363—364.
- Суханова, Бобылева, 2000, с. 384.
- Суханова, Бобылева, 2000, с. 383.
- Simpson AGB. Cytoskeletal organization, phylogenetic affinities and systematics in the contentious taxon Excavata (Eukaryota) // Int J Syst Evol Microbiol. — 2003. — Vol. 53. — P. 1759—1777. — doi:10.1099/ijs.0.02578-0.
- Суханова, Бобылева, 2000, с. 337.
- Хаусман и др., 2010, с. 67.
- Ruggiero M. A., Gordon D. P., Orrell T. M., Bailly N., Bourgoin T., Brusca R. C., Cavalier-Smith T., Guiry M. D., Kirk P. M. Correction: A Higher Level Classification of All Living Organisms // PLoS ONE, 2015, 10 (6): e0130114. — doi:10.1371/journal.pone.0130114
- Yao C., Köster L. S. Tritrichomonas foetus infection, a cause of chronic diarrhea in the domestic cat // Veterinary Research. — 2015. — Vol. 46. — P. 35. — doi:10.1186/s13567-015-0169-0.
- Hess M, Liebhart D, Bilic I, Ganas P. Histomonas meleagridis — new insights into an old pathogen // Vet Parasitol.. — 2015. — Vol. 208. — P. 67—76. — doi:10.1016/j.vetpar.2014.
- Parasites — Dientamoeba fragilis. Centers for Disease Control and Prevention.
- Ribeiro, L.C., Santos, C., Benchimol, M. Is Trichomonas tenax a Parasite or a Commensal? // Protist. — 2015. — Vol. 166. — P. 196—210. — doi:10.1016/j.protis.2015.02.002.
- Кисина В., Вавилов В., Гущин А. Урогенитальный трихомониаз: современный взгляд на проблему // Врач. — 2010. — № 1. — С. 18—20.
Литература
- Суханова К. М., Бобылева Н. Н. Polymastigota // Протисты = Protista : руководство по зоологии / гл. ред. А. Ф. Алимов. — СПб.: ЗИН РАН, 2000. — Т. 1. — С. 337—397.
- Хаусман К., Хюльсман H., Радек Р. Протистология / под. ред. С. Л. Корсуна. — Москва: Товарищество научных изданий КМК, 2010. — 495 с.
| Diaphore- tickes |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amorphea |
| ||||||||||||
| Положение не ясно |
| ||||||||||||
