Короткие открытые рамки считывания
Коро́ткие откры́тые ра́мки счи́тывания[1] (англ. upstream open reading frames, uORF) — открытые рамки считывания (англ. open reading frame, ORF), расположенные внутри 5′-нетранслируемой области (5'-UTR) эукариотических и некоторых вирусных[2] мРНК. uORF участвуют в регуляции экспрессии генов у эукариот и вирусов[3][4] и обычно подавляют трансляцию основной рамки считывания (то есть кодирующей последовательности), хотя их действие может сопровождаться различными эффектами[5].
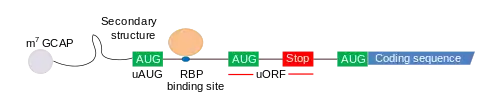
Функционирование
Механизмы, не зависящие от нуклеотидной последовательности uORF
В общем случае мРНК разделяется на 5’-нетранслируемый участок (5’-UTR), открытую рамку считывания (CDS), которая начинается со старт-кодона и заканчивается стоп-кодоном, а также 3-нетранслируемый участок (3’-UTR). Короткие открытые рамки считывания (uORF) находятся в 5’-UTR перед основной рамкой считывания. Впрочем, uORF может и перекрываться с основной кодирующей последовательностью (англ. coding sequence, CDS), тогда её стоп-кодон располагается после инициаторного кодона CDS[1].
uORF имеются в приблизительно 50 % 5’-UTR человеческих мРНК и по крайней мере 35 % мРНК млекопитающих[1], и их наличие обусловливает снижение экспрессии генов, уменьшая количество функциональной мРНК на 30 %, а образование белка — на 30—80 %. Рибосомы, связывающиеся со старт-кодоном короткой открытой рамки считывания (uAUG), начинают трансляцию uORF, что может отрицательно сказаться на эффективности трансляции основной рамки считывания (то есть кодирующей области). Если не происходит эффективного связывания рибосомы со старт-кодоном в кодирующей области (то есть инициации трансляции), то в результате снижается уровень образование белка, а значит, и уровень экспрессии соответствующего гена. Может произойти и обратная ситуация: трансляция uORF продолжится в трансляцию кодирующей области, и в итоге образуется слишком длинный белок, который может быть вредоносным для организма.
Снижение эффективности трансляции из-за наличия uORF в 5’-UTR является хорошо изученным эффектом; одним из примеров, иллюстрирующих его, может служить ген поли(А)-полимеразы α (англ. poly(A)-polymerase α, PAPOLA), чья мРНК содержит две высококонсервативные uORF в 5’-UTR. Мутация проксимального uAUG вызывает повышение эффективности трансляции этой мРНК, следовательно, uORF значительно снижает экспрессию этого гена. Другим примером является рецептор тиреоидного гормона, оказывающий активирующее или репрессирующее действие на транскрипцию ряда генов-мишеней: сильная репрессия его трансляции осуществляется uORF длиной 15 нуклеотидов внутри 5’-UTR его мРНК[6].
Широко распространена точка зрения, что uORF снижают эффективность трансляции, поскольку после терминации трансляции uORF рибосома не может вновь инициировать трансляцию и транслировать кодирующую область. Показано, что для успешной реинициации трансляции длина uORF не должна превышать 20 кодонов, однако удовлетворительного объяснения этого факта до сих пор не существует[1]. В ряде случаев и при определённых условиях старт-кодон uORF может быть проигнорирован сканирующей рибосомой. В то же время, под действием некоторых внутренних и внешних факторов ингибирующий эффект uORF может усиливаться, и главным из таких лимитирующих факторов является доступность инициаторного комплекса после терминации трансляции uORF[7].
Тем не менее, недавние исследования более чем 500 локусов генов, содержащих 5’-UTR, показали, что не существует определённой связи между влиянием uORF на экспрессию следующего за ней гена и расстоянием между uORF и кодирующей последовательностью. При этом авторы исследования предполагают, что в генах, содержащих единственную uORF, вероятнее всего, трансляция CDS осуществляется после сканирования uORF рибосомой без её диссоциации, а не через реинициацию трансляции. Это предположение сильно отличается от выводов Козак (1987) и вообще всех предшествующих представлений о uORF (тогда считалось верным во всех случаях правило, согласно которому рибосома начинает трансляцию с первого стартового кодона, который встретится ей при перемещении от 5'- к 3'-концу мРНК[8]).
Более того, эксперименты с клетками, лишёнными Rent1 (фактора, участвующего в процессе направленного разрушения дефектных мРНК — англ. nonsense-mediated decay, NMD), показали, что в отсутствие NMD успешно шла трансляция транскриптов, содержащих uORF. Это показывает, что NMD также играет важную роль в регуляции функционирования этих транскриптов. Вероятнее всего, существует несколько вариантов развития событий после взаимодействия uORF и рибосомы: продолжение трансляции, продолжение сканирования или реинициация трансляции кодирующей области, причём то, какой из них произойдёт, зависит от ряда факторов.
- GCN4
Так, мРНК гена GCN4 дрожжей, кодирующего активатор транскрипции, содержит 4 uORF в 5’-UTR, причём лишь одна из них позволяет рибосоме начать трансляцию кодирующей области. Эффективной реинициации способствуют два цис-энхансера, расположенные по обе стороны от uORF. Дальнейшие исследования показали, что 5’-цис-активирующая последовательность (энхансер) взаимодействует с N-концевым доменом субъединицы eIF3A/TIF32 фактора инициации eIF3, в результате чего 40S-субъединица рибосомы остаётся на мРНК после трансляции uORF1 и продолжает сканирование. Было также установлено, что реинициация облегчается специфическим сворачиванием мРНК во время осуществления сканирования рибосомой, и эта укладка позволяет взаимодействовать энхансеру с другой последовательностью мРНК. Это всего лишь один пример выключения гена с помощью uORF, однако к настоящему моменту очевидно, что механизм такого выключения гораздо более сложен, чем тот, который предполагает сканирующая модель. Для детального выяснения этого механизма или механизмов необходимы дальнейшие исследования[9].
- Протеинкиназа С
Ещё одним примером, иллюстрирующим сложность механизмов выключения гена, опосредованных uORF, может выступать протеинкиназа С (англ. protein kinase C, PKC). РКС является представителем семейства серин/треонин-протеинкиназ, участвующих в регуляции клеточного роста и дифференциации. Её новая изоформа, РКСη, тканеспецифична и экспрессируются в основном в быстро сменяющихся клетках, например, эпителиальных. Недавно было установлено, что эта изоформа играет особую роль в ответе на стресс, и её экспрессия коррелирует с устойчивостью к противораковым препаратам при ряде форм рака. 5’-UTR человеческой РКСη представляет собой длинную (659 нуклеотидов), GC-обогащённую последовательность и содержит 2 небольшие консервативные uORF. Мутации каждой из этих uORF вызывают небольшое увеличение экспрессии (в 1,5 и 2,2 раза) главной ORF, а мутация обеих — трёхкратное. Судя по всему, механизм репрессии трансляции РКСη имеет место в нормальных условиях. В условиях стресса (нехватке глюкозы или гипоксии) две uORF также задействованы в экспрессии, так как они обеспечивают слабое сканирование 5’-UTR и увеличивают трансляцию кодирующей последовательности. Варьирование в числе рибосом, связавшихся с транскриптом, и трансляции каждой uORF также могут обеспечивать клеткоспецифичную «подстройку» экспрессии генов[10].
- Орнитиндекарбоксилаза
Установлено, что, помимо AUG, в качестве сайта начала транскрипции могут использоваться также кодоны, отличающиеся от AUG одним нуклеотидом, и эффективность инициации в каждом случае будет определяться окружением нестандартного старт-кодона. Так, экспрессия орнитиндекарбоксилазы (англ. ornithine decarboxylase, ODC), участвующей в биосинтезе полиаминов, модулируется uORF со старт-кодоном АUU. Эффектиность инициации в этом случае варьирует в зависимости от внутриклеточной концентрации полиаминов, при этом эффективность инициации на AUU сильно снижена в клетках, обеднённых полиаминами: она составляет 18 % от эффективности трансляции основной ORF, а в клетках, насыщенных полиаминами, — 54 %. Таким образом поддерживается сокращенная экспрессия ODC при наличии полиаминов. Этот пример демонстрирует, что, вероятнее всего, существует гораздо большее число uORF, чем ранее предполагалось[10].
- Орнитиндекарбоксилаза
Хотя большая часть uORF отрицательно влияет на экспрессию генов, иногда наличие uORF усиливает трансляцию. Примером может служить бицистронная мРНК vpu-env вируса ВИЧ-1, содержащая консервативную очень маленькую uORF. Эта uORF располагается лишь за 5 нуклеотидов до AUG vpu и вскоре заканчивается стоп-кодоном, перекрывающимся с AUG vpu. Было установлено, что эта uORF оказывает значительный положительный эффект на трансляцию env и при этом не мешает трансляции vpu. Были получены мутанты, у которых расстояние между uORF и основным AUG было увеличено на 5 нуклеотидов, и было показано, что uORF не задействована в инициации vpu. На основании этого авторами исследования было высказано предположение, что эта маленькая uORF может служить местом задержки рибосомы, в ходе которой рибосома взаимодействует с РНК-структурами, способствующими её продвижению, то есть физически преодолевает часть 5’-UTR, чтобы достичь основного инициаторного кодона[11].
- РНКаза Н
За последнее десятилетие было показано, что регуляция экспрессии генов посредством uORF — сложный процесс. Хорошим примером такой сложной регуляции может служить следующий механизм. РНКаза Н1 имеется в ядрах и митохондриях клеток млекопитающих и по-разному экспрессируется в клетках разных типов. Экспрессия изоформ этого фермента находится под контролем двух внутрирамковых старт-кодонов AUG, а также uORF, локализованной в 5’-UTR. Трансляция РНКазы Н1 в митохондриях начинается с первого кодона AUG и обычно ограничивается uORF, в результате чего количество митохондриальной изоформы составляет около 10 % количества ядерной изоформы. Трансляция ядерной изоформы начинается со второго AUG и не зависит от uORF, поскольку рибосома способна эффективно реинициировать трансляцию со второго AUG, как бы перескакивая uORF. Такой механизм регуляции позволяет контролировать экспрессию РНКазы Н1 в митохондриях, где её избыток может привести к гибели клетки, и при этом не воздействовать на нормальный уровень экспрессии ядерной изоформы. Было также установлено, что изменение нуклеотидного окружения AUG вызывало накопление транскрипта, что свидетельствует об участии в этом механизме и других факторов. Этот пример иллюстрирует крайне специфичную систему регуляции экспрессии гена, где задействованы uORF и другие факторы[12].
Наконец, альтернативные промоторы и сплайсинг, а также то, что рибосома иногда может взаимодействовать с внерамковыми кодонами и использовать нестандартные старт-кодоны, даёт дополнительные возможности для регуляции экспрессии гена при участии uORF. Недавнее исследование в клеточной линии человеческих моноцитов, которая обрабатывалась пуромицином для преждевременной терминации трансляции и определялись сайты инициации трансляции, показало наличие 2994 новых uORF в одной только 5’-UTR, хотя, несомненно, многие uORF также перекрываются с кодирующей областью и 3’-UTR[13].
Механизмы, зависящие от нуклеотидной последовательности uORF
Собственно нуклеотидная последовательность uORF обычно не влияет на их действие; важны лишь длина, количество и расстояния между uORF[7]. Однако, в некоторых случаях эффект uORF всё-таки зависит от её нуклеотидной последовательности, в частности, от аминокислотной последовательности кодируемого ей пептида[14]. Хотя значение uORF как регуляторных элементов, участвующих в регуляции связывания рибосомы и трансляции, хорошо изучено, однако функция и даже судьба кодируемых uORF пептидов часто неизвестна, возможно, по причине сложностей в анализе уровня экспрессии и локализации пептидов. Доказательство того, что транслированные с uORF пептиды присутствует в клетках, было получено в 2004 году, кода было идентифицировано 54 пептида длиной менее 100 аминокислотных остатков. Эти пептиды образовывались в клетках хронической человеческой миелоидной лейкемии, для каждой из которых было установлено наличие uORF. Хотя эти пептиды были идентифицированы, для тысяч uORF в этих клетках неизвестно образование белков, которые можно идентифицировать экспериментально. Это может свидетельствать о том, что, во-первых, белки, транслируемые с uORF, могут селективно подвергаться протеолизу; во-вторых, некоторые uORF экспрессируются, но не в клетках этого типа; в-третьих, многие uORF не дают начала белкам. Впрочем, очевидно, что некоторые uORF всё же транслируются в пептиды, которые накапливаются в клетках и поэтому, видимо, несут функциональную нагрузку, хотя для многих ещё не установленную[12].
Тем не менее, в некоторых случаях механизм действия кодируемых uORF пептидов известен. Такой пептид функционирует как цис-регуляторный элемент и остаётся связанным с транслирующей рибосомой. По-видимому, механизм его действия связан со специфическим взаимодействием пептида с рибосомой, следствием чего является задержка терминации и невозможность дальнейшего продвижения рибосомы по мРНК. Примерами мРНК, чьи uORF кодируют ингибиторные пептиды, являются мРНК S-аденозилметиониндекарбоксилазы млекопитающих, мРНК CPA1 дрожжей, кодирующая фермент биосинтеза аргинина; мРНК gp48 цитомегаловируса человека. Длина этих пептидов составляет от 6 до 25 аминокислотных остатков, и было доказано, что именно их последовательность аминокислот обусловливает ингибиторный эффект[15].
Другие механизмы регуляции
Ингибирование трансляции кодирующей последовательности, обусловленное uORF, может регулироваться рядом транс-факторов и условиями окружающей среды. Например, трансляция уже упоминавшейся мРНК S-аденозилметиониндекарбоксилазы ингибируется uORF в покоящихся Т-клетках, однако трансляция успешно идёт в стимулированных Т-клетках и клетках Т-линии; в их случае, по-видимому, имеет место игнорирование uORF сканирующими рибосомами. Другой пример — также уже упоминавшаяся мРНК CPA1 дрожжей. В её случае uORF блокирует трансляцию основной рамки считывания только в присутствии аргинина. Точный механизм этой регуляции не установлен, но ингибирование пептидилтрансферазной функции в ответ на добавление аргинина останавливает рибосомы на стоп-кодоне uORF. Предполагается, что аргинин либо напрямую ингибирует пептидилтрансферазную активность, либо уменьшает доступность А-сайта рибосомы[16].
Интересно явление наблюдается при трансляции полицистронной РНК вируса мозаики цветной капусты (англ. cauliflower mosaic virus, CaMV). В её 5'-UTR имеются uORF, которые, как и предполагалось, оказывают подавляющее действие на трансляцию кодирующей последовательности. Более того, почти вся 5'-UTR укладывается в шпильку с развитой вторичной и третичной структурой, создающую преграду для передвижения рибосомы. Так и происходит в клетках растений, не являющихся хозяевами вируса, и в бесклеточных системах, приготовленных из их экстрактов. У растений-хозяев вируса трансляция, тем не менее, идёт, по-видимому, из-за наличия специального клеточного фактора и первых uORF. При подробном изучении выяснилось, что, когда РНК CaMV трансляционно активна, рибосомы пропускают центральную часть шпильки с пространственной структурой, как бы перепрыгивая её. Иными словами, в этом случае имеет место шунтирование, при котором рибосома считывает 3 первые uORF, при этом происходит расплетание основания шпильки, а рибосома входит в зону с такой структурой, что она может перепрыгнуть с участка сразу за третьей uORF на участок внутри седьмой uORF; после этого рибосома движется далее в направлении 3'-конца и начинает транслировать кодирующую область[17].
Клиническое значение
Мутации, затрагивающие uORF, как правило, вредны, поскольку они разрушают систему регуляции экспрессии гена, что может непосредственно привести к болезни. Например, мутации, разрушающие uORF в 5’-UTR гена, кодирующего белок HR (англ. human hairless homolog), приводят к аутосомно-доминантной форме облысения. Не менее вредны мутации, создающие новые uORF, поскольку они также нарушают нормальную регуляцию экспрессии гена. Было высказано предположение, что мутация в гене-супрессоре опухоли может привести к сниженной экспрессии защитных белков и развитию рака. Мутации в гене CDKN2A, кодирующем ингибитор протеинкиназ, могут стать предпосылками развития меланомы. Установлено, что мутации в uORF могут также приводить к развитию таких заболеваний, как наследственная тромбоцитемия, болезнь Альцгеймера, биполярное аффективное расстройство, кардиомиопатия, аритмогенная дисплазия правого желудочка[18]. Все эти примеры ещё раз доказывают исключительную важность uORF в тонкой регуляции экспрессии гена и поддержании гомеостаза, а вариабельность в участке uORF может обусловливать индивидуальный фенотип или предрасположенность к заболеваниям[19].
Примечания
- Спирин, 2011, с. 406.
- Спирин, 2011, с. 411.
- Vilela C., McCarthy J. E. Regulation of fungal gene expression via short open reading frames in the mRNA 5'untranslated region (англ.) // Microbiology : journal. — Microbiology Society, 2003. — August (vol. 49, no. 4). — P. 859—867. — doi:10.1046/j.1365-2958.2003.03622.x. — PMID 12890013.
- Lovett P. S., Rogers E. J. Ribosome regulation by the nascent peptide (англ.) // Microbiology and Molecular Biology Reviews : journal. — American Society for Microbiology, 1996. — June (vol. 60, no. 2). — P. 366—385. — PMID 8801438.
- Barrett et al. Untranslated Gene Regions and Other Non-coding Elements. — Basel: Springer, 2013. — С. 16—19. — 56 с. — ISBN 978-3-0348-0678-7.
- Barrett et al., 2013, p. 16.
- Спирин, 2011, с. 407.
- Mignone F., Gissi C., Liuni S., Pesole G. Untranslated regions of mRNAs. (англ.) // Genome biology. — 2002. — Vol. 3, no. 3. — P. 0004. — PMID 11897027.
- Barrett et al., 2013, p. 16—17.
- Barrett et al., 2013, p. 17.
- Barrett et al., 2013, p. 17—18.
- Barrett et al., 2013, p. 18.
- Barrett et al., 2013, p. 18—19.
- Спирин, 2011, с. 410.
- Спирин, 2011, с. 410—411.
- Wei J., Wu C., Sachs M. S. The arginine attenuator peptide interferes with the ribosome peptidyl transferase center. (англ.) // Molecular and cellular biology. — 2012. — Vol. 32, no. 13. — P. 2396—2406. — doi:10.1128/MCB.00136-12. — PMID 22508989.
- Спирин, 2011, с. 411—412.
- Sangeeta Chatterjee, Jayanta K. Pal. Role of 5- and 3-untranslated regions of mRNAs in human diseases // Biol. Cell. — 2009. — С. 251—262. — doi:10.1042/BC20080104. (недоступная ссылка)
- Barrett et al., 2013, p. 19.
Литература
- Спирин А. С. Молекулярная биология. Рибосомы и биосинтез белка. — М.: Издательский центр «Академия», 2011. — 496 с. — ISBN 978-5-7695-6668-4.
- Lucy W. Barrett, Sue Fletcher, Steve D. Wilton. Untranslated Gene Regions and Other Non-coding Elements. — SpringerBriefs in Biochemistry and Molecular Biology, 2013. — 57 p. — ISBN 978-3-0348-0679-4.