Биологические правила
Биологическое правило или биологический закон — это обобщенный закон, принцип или эмпирическое правило, сформулированное для описания закономерностей, наблюдаемых в живых организмах. Биологические правила и законы часто разрабатываются как краткие, широко применимые способы объяснения сложных явлений или важных наблюдений об экологии и биогеографическом распределении видов растений и животных по всему миру, хотя они были предложены или распространены для всех типов организмов. Многие из этих закономерностей экологии и биогеографии названы в честь биологов, впервые описавших их.[1][2]
С момента зарождения своей науки биологи стремились объяснить очевидные закономерности в данных наблюдениях. В своей биологии Аристотель вывел правила, регулирующие различия между живородящими четвероногими (в современных терминах — наземными плацентарными млекопитающими). Среди его правил было то, что размер выводка уменьшается с увеличением массы тела взрослой особи, в то время как продолжительность жизни увеличивается с периодом беременности и с массой тела, а плодовитость уменьшается с увеличением продолжительности жизни. Так, например, у слонов меньше выводков, чем у мышей, но при этом продолжительность жизни и беременности больше.[3] Подобные правила кратко систематизировали сумму знаний, полученных в результате ранних научных измерений мира природы, и могли использоваться в качестве моделей для прогнозирования будущих наблюдений. К числу самых ранних биологических правил в наше время относятся правила Карла Эрнста фон Бэра (начиная с 1828 года) об эмбриональном развитии[4] и Константина Вильгельма Ламберта Глогера о пигментации животных (1833 год).[5] Среди биогеографов существует некоторый скептицизм относительно полезности общих правил. Например, J. C. Briggs в своей книге 1987 года Биогеография и тектоника плит комментирует, что хотя правила кладистики Вилли Хеннига «в целом были полезны», его правило прогрессии «подозрительно».[6]
Список биологических правил
- Правило Аллена гласит, что среди родственных форм гомойотермных (теплокровных) животных, ведущих сходный образ жизни, те, которые обитают в более холодном климате, имеют относительно меньшие выступающие части тела: уши, ноги, хвосты и т. д. Он назван в честь Джоэла Азафа Аллена, который описал его в 1877 году.[8][9]
- Правило Бэтсона гласит, что дополнительные ноги зеркально-симметричны по отношению к своим соседям, например, когда дополнительная нога появляется в гнезде ноги насекомого. Он назван в честь пионера генетики Уильяма Бэтсона, который наблюдал его в 1894 году. Это, по-видимому, вызвано утечкой позиционных сигналов через границу раздела конечностей, так что полярность дополнительной конечности должна быть обратной.[10]
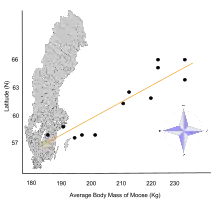
- Правило Бергмана гласит, что в пределах широко распространенной таксономической клады популяции и виды большего размера встречаются в более холодных средах, а виды меньшего размера — в более теплых регионах. Это относится, за исключением, ко многим млекопитающим и птицам. Он был назван в честь Карла Бергмана, который описал его в 1847 году.[11][12][13][14][15]
- Правило Копа гласит, что линии популяции животных имеют тенденцию увеличиваться в размерах тела в течение эволюционного времени. Правило названо в честь палеонтолога Эдварда Копа.[16][17]
- Глубоководный гигантизм, отмеченный в 1880 году Генри Ноттиджем Мозли,[18] утверждает, что глубоководные животные крупнее своих мелководных собратьев. В случае морских ракообразных было высказано предположение, что увеличение размера с глубиной происходит по той же причине, что и увеличение размера с широтой (правило Бергмана): обе тенденции включают увеличение размера с уменьшением температуры.[19]
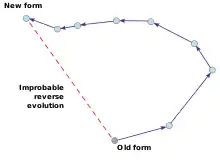 Закон необратимости Долло утверждает, что после того, как организм эволюционировал определенным образом, он не вернется точно к предыдущей форме.
Закон необратимости Долло утверждает, что после того, как организм эволюционировал определенным образом, он не вернется точно к предыдущей форме. - Закон необратимости Долло, предложенный в 1893 году[20] бельгийским палеонтологом французского происхождения Луи Долло, утверждает, что «организм никогда не возвращается точно в прежнее состояние, даже если он оказывается в условиях существования, идентичных тем, в которых он жил ранее … он всегда сохраняет некоторый след промежуточных стадий, через которые он прошел.»[21][22][23]
- Правило Эйхлера гласит, что таксономическое разнообразие паразитов колеблется вместе с разнообразием их хозяев. Он наблюдался в 1942 году Вольфдитрихом Эйхлером и назван в его честь.[24][25][26]
 Правило Эмери гласит, что насекомые- социальные паразиты такие как шмели-кукушки выбирают близкородственных хозяев, в данном случае других шмелей.
Правило Эмери гласит, что насекомые- социальные паразиты такие как шмели-кукушки выбирают близкородственных хозяев, в данном случае других шмелей. - Правило Эмери, отмеченное Карло Эмери, гласит, что социальные паразиты насекомых часто тесно связаны со своими хозяевами, например, принадлежат к одному роду.[27][28]
- Правило Фостера, также известное как правило острова или эффект острова гласит, что представители вида становятся меньше или больше в зависимости от ресурсов, доступных в окружающей среде.[29][30][31] Впервые это правило было сформулировано Дж. Бристолом Фостером в 1964 году в журнале Nature, в статье под названием «Эволюция млекопитающих на островах».[32]
- Закон Гаузе или принцип конкурентного исключения, названный в честь Георгия Гаузе, гласит, что два вида, конкурирующие за один и тот же ресурс, не могут сосуществовать при постоянных значениях численности популяции. Конкуренция ведет либо к вымиранию более слабого конкурента, либо к эволюционному или поведенческому сдвигу в сторону другой экологической ниши.[33]
- Правило Глогера гласит, что внутри вида эндотерм более сильно пигментированные формы, как правило, встречаются в более влажных средах, например вблизи экватора. Он был назван в честь зоолога Константина Вильгельма Ламберта Глогера, который описал его в 1833 году.[5][34]
- Правило Холдейна гласит, что если у гибридного вида бесплоден только один пол, то этот пол обычно является гетерогаметным. Гетерогаметный пол — это пол с двумя разными половыми хромосомами; у млекопитающих это самец с хромосомами XY. Правило названо в честь Джона Холдейна.[35]
- Правило Гамильтона гласит, что частота генов должна увеличиваться, когда родство реципиента с субъектом, умноженное на выгоду для реципиента, превышает репродуктивные издержки для субъекта. Это прогноз из теории родственного отбора сформулированный Уильямом Гамильтоном.[36]
- Правило Харрисона гласит, что размеры тела паразита изменяются вместе с размерами тела его хозяина. Он предложил это правило для вшей,[37] но более поздние авторы показали, что оно одинаково хорошо работает для многих других групп паразитов, включая ракушек, нематод,[38][39] блох, мух и клещей, а также для аналогичного случая мелких травоядных на больших растениях.[40][41][42]
- Правило прогрессии Хеннига гласит, что при рассмотрении группы видов в кладистике, виды с наиболее примитивными признаками обнаруживаются в самой ранней части области, которая будет центром происхождения этой группы. Названо в честь Вилли Хеннига, который изобрел это правило.[6][43]
- Правило Джордана гласит, что существует обратная зависимость между температурой воды и меристическими характеристиками, такими как количество лучей плавника, позвонков или числа чешуек, которые, как видно, увеличиваются с понижением температуры. Он назван в честь отца американской ихтиологии Дэвида Старра Джордана.[44]
 Принцип Лака соответствует размеру кладки для наибольшего количества детенышей, которых родители могут прокормить
Принцип Лака соответствует размеру кладки для наибольшего количества детенышей, которых родители могут прокормить - Принцип Лака, предложенный Дэвидом Лаком, гласит, что «размер кладок каждого вида птиц был адаптирован естественным отбором, чтобы соответствовать наибольшему количеству птенцов, для которых родители в среднем могут обеспечить достаточно пищи».[45]
- Правило Рапопорта гласит, что широтные диапазоны растений и животных обычно меньше на более низких широтах, чем на более высоких широтах. Он был назван в честь Эдуардо Х. Рапопорта Г. К. Стивенсом в 1989 году.[46]
- Правило Ренча гласит, что среди многих родственных видов половой диморфизм по размеру будет увеличиваться с увеличением размеров тела, когда самцы больше самок, и уменьшаться с увеличением средних размеров тела, когда самки больше самцов. Это правило применимо к приматам, ластоногим (тюленям), и даже к парнокопытным (таким как крупный рогатый скот и олени).[47] Он назван в честь Бернхарда Ренча, который предложил его в 1950 году.[48]
- Закон Шмальгаузена, названный в честь Ивана Шмальгаузена, гласит, что популяция находящаяся на крайнем пределе своей толерантности в каком-либо одном аспекте, более уязвима для небольших различий в любом другом аспекте. Следовательно, разброс данных — это не просто шум, мешающий обнаружению так называемых «основных эффектов», но также индикатор стрессовых условий, ведущих к большей уязвимости.[49]
- Правило Торсона гласит, что донные морские беспозвоночные в низких широтах имеют тенденцию производить большое количество яиц, развивающихся до пелагических (часто планктотрофных — питающихся планктоном) и широко рассеивающихся личинок, тогда как в высоких широтах такие организмы имеют тенденцию производить меньше и больше лецитрофных (питающихся желтком) яиц и более крупных потомков, часто живородящих или яйцеживородящих, которые чаще вынашиваются.[50] Названо в честь Гуннара Торсона С. А. Милейковским в 1971 году.[51]
- Закон Ван Валена гласит, что вероятность исчезновения видов и высших таксонов (таких как семейства и отряды) постоянна для каждой группы с течением времени; группы не становятся ни более устойчивыми, ни более уязвимыми к вымиранию, независимо от возраста их родословной. Он назван в честь биолога-эволюциониста Ли Ван Валена.[52]
- Законы фон Бэра, открытые Карлом Эрнстом фон Бэром, гласят, что эмбрионы исходят из общей формы и развиваются во все более специализированные формы, так что разнообразие эмбриональных форм отражает таксономическое и филогенетическое древо. Таким образом, все животные в типе имеют сходный ранний эмбрион; животные в более мелких таксонах (классах, отрядах, семействах, родах, видах) имеют более поздние и поздние эмбриональные стадии. Это резко контрастировало с теорией рекапитуляции Иоганна Фридриха Меккеля (а позднее и Эрнста Геккеля), которая утверждала, что эмбрионы проходят через стадии, напоминающие взрослые организмы, от последовательных стадий scala naturae от предположительно низших до высших уровней организации.[53][54][4]
- Закон Уиллистона, впервые замеченный Сэмюэлем Уиллистоном, гласит, что части организма имеют тенденцию уменьшаться в количестве и значительно специализироваться в функциях. Он изучал зубные ряды позвоночных и заметил, что там, где у древних животных были рты с различными видами зубов, у современных хищников появились резцы и клыки, предназначенные для разрывания и разрезания плоти, в то время как у современных травоядных появились большие коренные зубы, предназначенные для измельчения твердых растительных материалов.[55]
Ссылки
- Jørgensen, Sven Erik (2002). “Explanation of ecological rules and observation by application of ecosystem theory and ecological models”. Ecological Modelling. 158 (3): 241—248. DOI:10.1016/S0304-3800(02)00236-3.
- Allee, W. C. Ecological Animal Geography / W. C. Allee, K. P. Schmidt. — 2nd. — Joh Wiley & sons, 1951. — P. 457, 460–472.
- Leroi, Armand Marie. The Lagoon: How Aristotle Invented Science. — Bloomsbury, 2014. — P. 408. — ISBN 978-1-4088-3622-4.
- Lovtrup, Soren (1978). “On von Baerian and Haeckelian Recapitulation”. Systematic Zoology. 27 (3): 348—352. DOI:10.2307/2412887. JSTOR 2412887.
- Gloger, Constantin Wilhelm Lambert. 5. Abänderungsweise der einzelnen, einer Veränderung durch das Klima unterworfenen Farben // Das Abändern der Vögel durch Einfluss des Klimas : []. — Breslau : August Schulz, 1833. — P. 11–24. — ISBN 978-3-8364-2744-9.
- Briggs, J. C. Biogeography and Plate Tectonics. — Elsevier, 1987. — P. 11. — ISBN 978-0-08-086851-6.
- Sand, Håkan K.; Cederlund, Göran R.; Danell, Kjell (June 1995). “Geographical and latitudinal variation in growth patterns and adult body size of Swedish moose (Alces alces)”. Oecologia. 102 (4): 433—442. Bibcode:1995Oecol.102..433S. DOI:10.1007/BF00341355. PMID 28306886.
- Allen, Joel Asaph (1877). “The influence of Physical conditions in the genesis of species”. Radical Review. 1: 108—140.
- Lopez, Barry Holstun. Arctic Dreams: Imagination and Desire in a Northern Landscape. — Scribner, 1986. — ISBN 978-0-684-18578-1.
- Held, Lewis I.; Sessions, Stanley K. (2019). “Reflections on Bateson's rule: Solving an old riddle about why extra legs are mirror‐symmetric”. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution. 332 (7): 219—237. DOI:10.1002/jez.b.22910. ISSN 1552-5007. PMID 31613418.
- Olalla-Tárraga, Miguel Á.; Rodríguez, Miguel Á.; Hawkins, Bradford A. (2006). “Broad-scale patterns of body size in squamate reptiles of Europe and North America”. Journal of Biogeography. 33 (5): 781—793. DOI:10.1111/j.1365-2699.2006.01435.x.
- Timofeev, S. F. (2001). “Bergmann's Principle and Deep-Water Gigantism in Marine Crustaceans”. Biology Bulletin (Russian Version, Izvestiya Akademii Nauk, Seriya Biologicheskaya). 28 (6): 646–650 (Russian version, 764–768). DOI:10.1023/A:1012336823275.
- Meiri, S.; Dayan, T. (2003-03-20). “On the validity of Bergmann's rule”. Journal of Biogeography. 30 (3): 331—351. DOI:10.1046/j.1365-2699.2003.00837.x.
- Ashton, Kyle G.; Tracy, Mark C.; Queiroz, Alan de (October 2000). “Is Bergmann's Rule Valid for Mammals?”. The American Naturalist. 156 (4): 390—415. DOI:10.1086/303400. JSTOR 10.1086/303400. PMID 29592141.
- Millien, Virginie; Lyons, S. Kathleen; Olson, Link; et al. (May 23, 2006). “Ecotypic variation in the context of global climate change: Revisiting the rules”. Ecology Letters. 9 (7): 853—869. DOI:10.1111/j.1461-0248.2006.00928.x. PMID 16796576.
- Rensch, B. (September 1948). “Histological Changes Correlated with Evolutionary Changes of Body Size”. Evolution. 2 (3): 218—230. DOI:10.2307/2405381. JSTOR 2405381.
- Stanley, S. M. (March 1973). “An Explanation for Cope's Rule”. Evolution. 27 (1): 1—26. DOI:10.2307/2407115. JSTOR 2407115.
- McClain, Craig. Why isn't the Giant Isopod larger?. Deep Sea News (14 января 2015).
- Timofeev, S. F. (2001). “Bergmann's Principle and Deep-Water Gigantism in Marine Crustaceans”. Biology Bulletin (Russian Version, Izvestiya Akademii Nauk, Seriya Biologicheskaya). 28 (6): 646–650 (Russian version, 764–768). DOI:10.1023/A:1012336823275.
- Dollo, Louis (1893). “Les lois de l'évolution” (PDF). Bull. Soc. Belge Geol. Pal. Hydr. VII: 164—166.
- Gould, Stephen J. (1970). “Dollo on Dollo's law: irreversibility and the status of evolutionary laws”. Journal of the History of Biology. 3 (2): 189—212. DOI:10.1007/BF00137351. PMID 11609651.
- Goldberg, Emma E.; Boris Igić (2008). “On phylogenetic tests of irreversible evolution”. Evolution. 62 (11): 2727—2741. DOI:10.1111/j.1558-5646.2008.00505.x. PMID 18764918.
- Collin, Rachel; Maria Pia Miglietta (2008). “Reversing opinions on Dollo's Law”. Trends in Ecology & Evolution. 23 (11): 602—609. DOI:10.1016/j.tree.2008.06.013. PMID 18814933.
- Eichler, Wolfdietrich (1942). “Die Entfaltungsregel und andere Gesetzmäßigkeiten in den parasitogenetischen Beziehungen der Mallophagen und anderer ständiger Parasiten zu ihren Wirten” (PDF). Zoologischer Anzeiger. 136: 77—83.
- Klassen, G. J. (1992). “Coevolution: a history of the macroevolutionary approach to studying host-parasite associations”. Journal of Parasitology. 78 (4): 573—87. DOI:10.2307/3283532. JSTOR 3283532. PMID 1635016.
- Vas, Z.; Csorba, G.; Rozsa, L. (2012). “Evolutionary co-variation of host and parasite diversity – the first test of Eichler's rule using parasitic lice (Insecta: Phthiraptera)” (PDF). Parasitology Research. 111 (1): 393—401. DOI:10.1007/s00436-012-2850-9. PMID 22350674.
- Richard Deslippe. Social Parasitism in Ants. Nature Education Knowledge (2010). — «В 1909 году систематик Карло Эмери сделал важное обобщение, известное теперь как правило Эмери, которое гласит, что социальные паразиты и их хозяева имеют общее происхождение и, следовательно, тесно связаны друг с другом (Эмери 1909).».
- Emery, Carlo (1909). “Über den Ursprung der dulotischen, parasitischen und myrmekophilen Ameisen”. Biologisches Centralblatt [нем.]. 29: 352—362.
- Juan Luis Arsuaga, Andy Klatt, The Neanderthal’s Necklace: In Search of the First Thinkers, Thunder’s Mouth Press, 2004, ISBN 1-56858-303-6, ISBN 978-1-56858-303-7, p. 199.
- Jean-Baptiste de Panafieu, Patrick Gries, Evolution, Seven Stories Press, 2007, ISBN 1-58322-784-9, ISBN 978-1-58322-784-8, p 42.
- Lomolino, Mark V. (February 1985). “Body Size of Mammals on Islands: The Island Rule Reexamined”. The American Naturalist. 125 (2): 310—316. DOI:10.1086/284343. JSTOR 2461638.
- Foster, J. B. (1964). “The evolution of mammals on islands”. Nature. 202 (4929): 234—235. Bibcode:1964Natur.202..234F. DOI:10.1038/202234a0.
- Hardin, Garrett (1960). “The competitive exclusion principle” (PDF). Science. 131 (3409): 1292—1297. Bibcode:1960Sci...131.1292H. DOI:10.1126/science.131.3409.1292. PMID 14399717.
- Zink, R. M.; Remsen, J. V. (1986). “Evolutionary processes and patterns of geographic variation in birds”. Current Ornithology. 4: 1—69.
- Turelli, M.; Orr, H. Allen (May 1995). “The Dominance Theory of Haldane's Rule”. Genetics. 140 (1): 389—402. PMC 1206564. PMID 7635302.
- Queller, D. C.; Strassman, J. E. (2002). “Quick Guide: Kin Selection” (PDF). Current Biology. 12 (24): R832. DOI:10.1016/s0960-9822(02)01344-1. PMID 12498698. Архивировано из оригинала (PDF) 2011-06-04.
- Harrison, Launcelot (1915). “Mallophaga from Apteryx, and their significance; with a note on the genus Rallicola” (PDF). Parasitology. 8: 88—100. DOI:10.1017/S0031182000010428. Архивировано из оригинала (PDF) 2017-11-07.
- Morand, S.; Legendre, P.; Gardner, SL; Hugot, JP (1996). “Body size evolution of oxyurid (Nematoda) parasites: the role of hosts”. Oecologia. 107 (2): 274—282. Bibcode:1996Oecol.107..274M. DOI:10.1007/BF00327912. PMID 28307314.
- Morand, S.; Sorci, G. (1998). “Determinants of life-history evolution in nematodes”. Parasitology Today. 14 (5): 193—196. DOI:10.1016/S0169-4758(98)01223-X. PMID 17040750.
- Harvey, P. H.; Keymer, A. E. (1991). “Comparing life histories using phylogenies”. Philosophical Transactions of the Royal Society B. 332 (1262): 31—39. Bibcode:1991RSPTB.332...31H. DOI:10.1098/rstb.1991.0030.
- Morand, S.; Hafner, M. S.; Page, R. D. M.; Reed, D. L. (2000). “Comparative body size relationships in pocket gophers and their chewing lice” (PDF). Zoological Journal of the Linnean Society. 70 (2): 239—249. DOI:10.1111/j.1095-8312.2000.tb00209.x. Архивировано из оригинала (PDF) 2018-03-31.
- Johnson, K. P.; Bush, S. E.; Clayton, D. H. (2005). “Correlated evolution of host and parasite body size: tests of Harrison's rule using birds and lice” (PDF). Evolution. 59 (8): 1744—1753. DOI:10.1111/j.0014-3820.2005.tb01823.x. Архивировано из оригинала (PDF) 2018-03-31.
- Centers of Origin, Vicariance Biogeography. The University of Arizona Geosciences.
- McDowall, R. M. (March 2008). “Jordan's and other ecogeographical rules, and the vertebral number in fishes”. Journal of Biogeography. 35 (3): 501—508. DOI:10.1111/j.1365-2699.2007.01823.x.
- Lack, David. The regulation of animal numbers. — Clarendon Press, 1954.
- Stevens, G. C. (1989). “The latitudinal gradients in geographical range: how so many species co-exist in the tropics”. American Naturalist. 133 (2): 240—256. DOI:10.1086/284913.
- Fairbairn, D. J. (1997). “Allometry for Sexual Size Dimorphism: Pattern and Process in the Coevolution of Body Size in Males and Females”. Annu. Rev. Ecol. Syst. 28 (1): 659—687. DOI:10.1146/annurev.ecolsys.28.1.659.
- Rensch, Bernhard (1950). “Die Abhängigkeit der relativen Sexualdifferenz von der Körpergrösse”. Bonner Zoologische Beiträge. 1: 58—69.
- Lewontin, Richard; Levins, Richard (2000). “Schmalhausen's Law”. Capitalism, Nature, Socialism. 11 (4): 103—108. DOI:10.1080/10455750009358943.
- Thorson, G. 1957 Донные сообщества (сублиторальные или мелководные шельфовые). В «Трактате по морской экологии и палеоэкологии» (Под ред. J.W. Hedgpeth) стр. 461—534. Геологическое общество Америки.
- Mileikovsky, S. A. 1971. Типы развития личинок у морских донных беспозвоночных, их распространение и экологическое значение: переоценка. Морская биология 19: 193—213
- Leigh Van Valen, evolutionary theorist and paleobiology pioneer, 1935-2010. University of Chicago.
- Opitz, John M.; Schultka, Rüdiger; Göbbel, Luminita (2006). “Meckel on developmental pathology”. American Journal of Medical Genetics Part A. 140A (2): 115—128. DOI:10.1002/ajmg.a.31043. PMID 16353245.
- Garstang, Walter (1922). “The Theory of Recapitulation: A Critical Re-statement of the Biogenetic Law”. Journal of the Linnean Society of London, Zoology. 35 (232): 81—101. DOI:10.1111/j.1096-3642.1922.tb00464.x.
- Williston, Samuel Wendall. Water Reptiles of the Past and Present. — Chicago : University of Chicago Press, 1914.