Фитоценоз
Фитоцено́з (от др.-греч. φυτóν «растение» + κοινός «общий») — растительное сообщество, существующее в пределах одного биотопа. Характеризуется относительной однородностью видового состава, определённой структурой и системой взаимоотношений растений друг с другом и со внешней средой. По Н. Баркману[1] фитоценоз — суть конкретный сегмент растительности, в котором внутренние флористические различия меньше, чем различия с окружающей растительностью. Термин предложен польским ботаником И. К. Пачоским в 1915 году. Фитоценозы является объектом изучения науки фитоценологии (геоботаники).

Фитоценоз входит в состав биоценоза наряду с зооценозом и микробиоценозом. Биоценоз, в свою очередь, в сочетании с условиями абиотический среды (экотопом) образует биогеоценоз. Фитоценоз является центральным, ведущим элементом биогеоценоза, так как трансформирует первичный экотоп в биотоп, создавая среду обитания для других организмов, а также является первым звеном в круговороте веществ и энергии. От растительности зависят свойства почв, микроклимат, состав животного мира, такие характеристики биогеоценоза, как биомасса, биопродуктивность и т. д. В свою очередь, элементами фитоценоза являются ценопопуляции растений — совокупности особей одного вида в границах фитоценозов.
Развитие взглядов на природу фитоценоза
На заре развития геоботаники оформилось представление о фитоценозе как реально существующей дискретной единице растительного покрова, что на тот момент представлялось вполне целесообразным, так как выделение отдельных фитоценозов существенно облегчало задачу изучения растительности в целом. Однако в начале XX века была высказана[2][3][4][5] диаметрально противоположная точка зрения, согласно которой растительный покров представлялся непрерывным, а его подразделение на отдельные элементы — фитоценозы — искусственным. Отсутствие резких границ между растительными сообществами и наличие переходных зон между ними способствовало возникновению учения о непрерывности (континуальности) растительного покрова, основанного на индивидуалистической концепции:
- Каждый вид растений индивидуален по своим требованиям к условиям внешней среды и имеет характерные экологические амплитуды по каждому экологическому фактору
- Факторы среды изменяются постепенно, как в пространстве, так и во времени
- Переход от одной комбинации ценопопуляций к другой осуществляется непрерывно: одни виды постепенно уменьшают своё обилие и исчезают, другие — появляются и увеличивают.
Крайними сторонниками концепции континуальности растительного покрова в качестве объекта изучения геоботаники рассматривался не фитоценоз, в силу его искусственности, а отдельное растение. Крайние сторонники идеи дискретности же постулировали чёткое различие и разграничение отдельных фитоценозов.
На основе синтеза обеих концепций была выдвинута[6][7] идея о сочетании в природе растительного покрова и дискретности, и непрерывности. Это представлялось как одно из проявлений противоречивости, свойственной для материального мира в целом. Согласно этой идеи, растительный покров обладает свойством непрерывности, но она не абсолютна, а относительна. В то же время он обладает и свойством дискретности, но и она не абсолютна, а относительна. Эти свойства органично сочетаются, не исключая, а дополняя друг друга.
Формирование фитоценоза

Формирование фитоценозов можно рассматривать как в динамическом аспекте (смена сообществ), так и в плане образования их на свободных участках земной поверхности.
Различают первично свободные участки, которые в прошлом не заселялись растениями и не содержат их зачатков. Фитоценозы на них могут образоваться только при внесении диаспор извне. К таким участкам относятся скальные обнажения, свежие речные и морские наносы, обнажившееся дно водоёмов, участки, освободившиеся от ледников, лавовые поля и др. В целом на Земле они занимают незначительные площади.
Вторично свободные участки образуются в местах, где раньше растительность существовала, но была уничтожена вследствие воздействия какого-либо неблагоприятного фактора. Примером могут служить гари, осыпи, незасеянные пашни, участки выеденных вредителями или скотом фитоценозов. На них в большинстве случаев сохраняется почва и диаспоры, а формирование фитоценозов происходит значительно быстрее, чем на первично свободных участках.
Формирование фитоценоза является непрерывным процессом, но условно может быть разделено на стадии:
- Отсутствие фитоценоза — случайный состав видов; отсутствие взаимодействия между растениями; очень слабое влияние на среду; невыраженность структуры.
- Открытый фитоценоз — неустойчивый состав, преимущественно из однолетников; структура с отдельными не взаимодействующими друг с другом ценопопуляциями.
 Сгоревший лес — вторично свободный участок
Сгоревший лес — вторично свободный участок - Закрытый невыработавшийся фитоценоз — выпадение значительной части видов-первопоселенцев; пятнистая структура с проникновением отдельных растений в скопления других видов; намечается ярусность.
- Закрытый выработавшийся фитоценоз — относительно постоянный видовой состав; затруднение проникновения новых видов; взаимодействие всех ценопопуляций; выраженная ярусность.
- Пионерная группировка — ценопопуляции малочисленны, взаимоотношения между ними отсутствуют
- Группово-зарослевое сообщество — ценопопуляции распределены куртинами, в которых происходит взаимодействие между растениями
- Диффузное сообщество — ценопопуляции смешиваются, вырабатывается система межвидовых взаимодействий
- по Ф. Клементсу[10]:
- Миграция — занос диаспор
- Эцезис — закрепление первопоселенцев
- Агрегация — образование групп потомства вокруг материнских растений
- Инвазия — перемешивание ценопопуляций
- Конкуренция — выработка конкурентных отношений вследствие резкого увеличения сомкнутости
- Стабилизация — формирование устойчивого замкнутого сообщества
Е. П. Прокопьев, суммируя различные схемы членения процесса формирования фитоценоза, предлагает выделять в нём три этапа[11]:
- Поступление зачатков на свободный участок. Видовой состав формирующегося фитоценоза будет зависеть от видового состава растений окружающей территории и характера распространения их диаспор, причём основную роль будут играть зачатки аллохорических видов, главным образом анемохоры.
- Экотопический (абиотический) отбор. Не все попавшие на свободный участок диаспоры на нём приживутся: часть не прорастёт, а часть проросших погибнет в молодом состоянии вследствие неблагоприятного сочетания абиотических факторов. Закрепившиеся растения будут являться пионерными для данной территории.
- Фитоценотический отбор. Вследствие размножения и расселения пионерных видов по участку, они начнут влиять друг на друга и изменять экотоп, формируя биотоп (местообитание). Первичная абиотическая среда экотопа превращается во вторичную биотическую — фитосреду. Под влиянием фитосреды и взаимовлияний растений некоторые пионерные виды, не приспособленные к ней, выпадают. Это может происходить, например, вследствие затенения или аллелопатии. Параллельно происходит закрепление на участке новых видов, уже приспособленных к данной фитосреде.
Факторы организации фитоценоза
Факторы организации растительного сообщества можно условно разделить на четыре группы: характеристики среды (экотопа), взаимоотношение между растениями, влияние на растительность гетеротрофных компонентов (животных, грибов, бактерий) и нарушения. Эти группы факторов определяют сочетание и характеристики ценопопуляций видов в фитоценозе.
Экотоп является главным фактором организации фитоценоза, хотя он может быть в значительной степени трансформирован биотическими влияниями растений или нарушениями. К абиотическим факторам, влияющим на организацию сообщества можно отнести:
- климатические (световой, тепловой, водный режимы и др.)
- эдафические (гранулометрический и химический состав, влажность, порозность, водный режим и другие свойства почв и грунтов)
- топографические (характеристики рельефа)
Взаимоотношения растений подразделяются на контактные и опосредованные: трансабиотические — через абиотические факторы среды обитания и трансбиотические — через третьи организмы.
Влияние на организацию фитоценозов гетеротрофных компонентов биогеоценозов исключительно разнообразно. Влияние животных проявляется в опылении, поедании, распространении семян, изменении стволов и крон деревьев и связанных с ними характеристик, разрыхлении почвы, вытаптывании и др. Микоризные грибы улучшают снабжение растений элементами минерального питания и водой, повышают устойчивость к патогенам. Бактерии-азотфиксаторы повышают снабжение растений азотом. Другие грибы и бактерии, а также вирусы могут являться патогенами.
Нарушения, как антропогенного, так и природного генезиса могут полностью трансформировать фитоценоз. Это происходит при пожарах, вырубках, выпасе скота, рекреационной нагрузке и т. п. В этих случаях формируются производные фитоценозы, которые постепенно изменяются в сторону восстановления коренного, если воздействие нарушающего агента прекратилось. Если воздействие долговременно (например, при рекреации) формируются сообщества, приспособленные к существованию при данном уровне нагрузки. Деятельность человека привела к образованию фитоценозов, ранее не существовавших в природе (например, сообществ на токсичных отвалах промышленных производств).
Взаимовлияния организмов в фитоценозах
Наличие системы взаимоотношений между растениями является одним из главных признаков сложившегося фитоценоза. Изучение их, в силу большой перекрываемости и сильного влияния абиотических факторов, представляет собой трудную задачу и может быть реализовано либо в виде эксперимента, в ходе которого изучаются взаимоотношения двух конкретных видов, либо вычленением таких отношений из комплекса других с применением методов математического анализа.
Прямые (контактные) взаимовлияния

Возникают при соприкосновении или проникновении организмов друг в друга. Подразделяются на физиологические (паразитизм и симбиоз), когда между организмами осуществляется активный обмен веществом и энергией, и механические (взаимоотношения эпифитов с форофитами и лиан с опорными растениями) — когда таковой отсутствует.
При паразитизме один организм (паразит) использует другой (хозяина) для получения необходимых ему веществ и энергии, при этом его угнетая. Среди высших растений паразитизм встречается только у покрытосеменных (см. растения-паразиты). Также на всех растениях способны поселяться паразитические грибы и бактерии. Степень поражения растения паразитами зависит от особенностей растения-хозяина (разные виды поражаются неодинаково), от условий местообитания (в условиях засоления растения практически не поражаются), от наличия эволюционной сопряжённости паразита и хозяина (если её нет, у хозяина обычно отсутствуют механизмы защиты от паразита).
Против паразитических грибов и бактерий у растений в ходе эволюции сформировался комплекс защитных механизмов:
- выделение растением фунгицидных и бактерицидных веществ, предотвращающих заражение паразитами или подавляющих их развитие
- наличие мощных покровных тканей, препятствующих проникновению паразитов
- особенности биохимического состава и метаболизма в клетках растения, препятствующие росту паразитов
Несмотря на наличие подобных механизмов защиты, фитопаразиты способны вызывать многочисленные болезни растений, ведущие к их ослаблению и гибели. Растения-паразиты распространены не столь широко, но также могут значительно угнетать ценопопуляции и отдельные растения.

Симбиотические отношения проявляются в сосуществовании растений с грибами и бактериями (в том числе цианобактериями). Соответственно различают микосимбиотрофию и бактериосимбиотрофию.
Микосимбиотрофия реализуется в виде микоризы — взаимодействия гиф грибов и корней растений. Предполагается[11], что на ранних этапах эволюции растительного мира грибы выступали по отношению к растениям только как паразиты и лишь в процессе длительной коэволюции сформировались взаимовыгодные отношения. К настоящему времени микориза обнаружена более чем у 80 % видов сосудистых растений: её образуют все виды голосеменных, 77—78 % видов покрытосеменных и около 60 % видов сосудистых споровых растений[12]. Различают эктомикоризу, когда мицелий опутывает корни растения и эндомикоризу, когда проникает в них.
Экологический смысл образования микоризы состоит в том, что гриб получает от растения углеводы и некоторые витамины, а растение следующие выгоды:
- мицелий гриба увеличивает всасывающую поверхность корней растения, улучшая снабжение его водой и минеральными веществами
- гриб может выделять некоторые витамины и ростовые вещества
- разлагая недоступные растениям органические вещества, грибы преобразуют их в доступную усвояемую форму
- заражение корней микоризными грибами предохраняет растение от воздействия патогенных микроорганизмов, в том числе паразитических грибов
- при эндомикоризе часть грибных клеток в корнях растений разлагается и используется в качестве источника питательных веществ
Растения, образующие микоризу, по требовательности к наличию микосимбионта можно разделить на две группы:
- облигатные микосимбиотрофы — не способные к развитию без микосибионта (сем. Орхидные)
- факультативные микосимбиотрофы — способные к существованию без микосимбионта, но лучше развивающиеся при его наличии
Бактериосимбиотрофия — симбиоз растений с клубеньковыми бактериями (Rhizobium sp.). Распространена не столь широко, как микосимбиотрофия — в симбиоз с бактериями вступает порядка 3 % растений мировой флоры (в основном семейства Бобовые (около 86 % видов семейства), а также некоторые виды семейств Мятликовые, Берёзовые, Лоховые, Крушиновые)[12][13]. Клубеньковые бактерии играют роль азотфиксаторов, переводя атмосферный азот в доступные растениям формы. Выделяют корневую и листовую формы взаимодействия. При корневой форме бактерии заражают корни растения, вызывая интенсивные локальные деления клеток и образование клубеньков. Листовая бактериосимбиотрофия встречается у некоторых тропических растений и ещё слабо изучена.
На способности растений семейства Бобовых вступать в симбиоз с клубеньковыми бактериями основан один из методов трёхпольной культуры.

Эпифиты, поселяющиеся на растениях-форофитах, используют последние только в качестве субстрата, не вступая с ними в физиологические взаимодействия. Эпифитные формы встречаются в группах покрытосеменных, папоротников, мхов, водорослей и лишайников. Наибольшего разнообразия эпифиты достигают во влажных тропических лесах.
В экологическом плане взаимоотношения эпифитов и форофитов обычно представлены комменсализмом, но могут проявляться и элементы конкуренции:
- эпифиты частично перехватывают свет и влагу у форофитов
- задерживая влагу, способствуют загниванию форофита
- затеняя форофит, эпифиты уменьшают его эффективную фотосинтетическую поверхность
- обильно разрастаясь, могут быть причиной деформации или слома форофитов
Лианы — растения, использующие другие растения или иные объекты для сохранения вертикального положения. Лианы появились в ходе эволюции как одно из проявлений конкуренции за свет. Подавляющее число видов лиан относится к покрытосеменным растениям. Большинство лиан укореняется в почве, однако существуют эпифитные и паразитные формы, например, растения рода Повилика (Cuscuta) сем. Вьюнковые (Convolvulaceae).
Лианы, вынося в ходе роста свои листья ближе к свету, получают от сожительства с опорным растением пользу, тогда как последнее — преимущественно вред, как прямой — вследствие механического воздействия лианы и слома / гибели опорного растения[14], так и косвенный — вследствие перехвата лианой света, влаги и питательных веществ.
Наибольшего разнообразия лианы также достигают во влажных тропических лесах.
Трансабиотические взаимовлияния
Влияния растений друг на друга, опосредованные абиотическими факторами внешней среды. Возникают благодаря перекрыванию фитогенных полей расположенных по соседству растений. Подразделяются на конкуренцию и аллелопатию.
Конкуренция развивается либо из-за исходной ограниченности ресурсов местообитания, либо в результате уменьшения их доли, приходящейся на каждое растение, вследствие перенаселённости. Конкуренция ведёт к снижению потребления ресурсов растением и, как следствие, уменьшению скорости роста и запасания веществ, а это, в свою очередь, приводит к снижению количества и качества диаспор. Различают внутри- и межвидовую конкуренцию.
Внутривидовая конкуренция влияет на показатели рождаемости и смертности в ценопопуляции, определяя тенденцию к поддержанию её численности на определённом уровне, когда обе величины уравновешивают друг друга. Такая численность называется предельной плотностью и зависит от количества ресурсов местообитания. Внутривидовая конкуренция асимметрична — неодинаково влияет на разные особи. Суммарная фитомасса ценопопуляции остаётся постоянной в достаточно большом диапазоне значений плотности, тогда как средняя масса одного растения при загущении начинает неуклонно снижаться — закон постоянства урожая (C=dw, где C — урожай, d — плотность ценопопуляции и w — средняя масса одного растения).
Межвидовая конкуренция также широко распространена в природе, поскольку абсолютное большинство фитоценозов (кроме некоторых агроценозов) являются многовидовыми. Многовидовой состав обеспечивается тем, что каждый вид имеет характерную только для него экологическую нишу, которую и занимает в сообществе. При этом ниша, которую вид мог бы занять при отсутствии межвидовой конкуренции — фундаментальная, сужается до размеров реализованной. В фитоценозе дифференциация экологических ниш происходит за счёт:
- разной высоты растений
- разной глубины проникновения корневой системы
- контагиозного распределения особей популяции (отдельными группами / пятнами)
- разных сроков вегетации, цветения и плодоношения
- неравной эффективности использования растениями ресурсов местообитания
При слабом перекрывании экологических ниш может наблюдаться сосуществование двух ценопопуляций, при сильном же более конкурентоспособный вид вытесняет из местообитания менее конкурентоспособный. Сосуществование двух сильно конкурирующих видов также возможно за счёт динамичности среды, когда тот или иной вид получает временное преимущество.
Аллелопатия — влияние растений друг на друга и на другие организмы путём выделения в среду активных метаболитов как при жизни растения, так и при разложении его остатков. Аллелопатическая активность того или иного вида обуславливается определённым набором химических веществ различной природы, качественный и количественный состав которого существенно зависит от внешних условий. Аллелопатически активные вещества выделяются как надземными органами (главным образом листьями), так и подземными в основном тремя способами:
- активное выделение через железы или гидатоды
- вымывание атмосферными осадками
- выделение посредством разложения опада микроорганизмами
Сумма выделений различных растений в фитоценозе — его биохимическая среда. Так как состав выделений не постоянен, можно говорить о существовании аллелопатического режима фитоценоза, наряду с водным, воздушным и др.
Трансбиотические взаимовлияния
Косвенные влияния одних растений на другие посредством третьих организмов (других растений, животных или грибов). Воздействие может проявляться как на уровне отдельного организма, так и на уровне целой ценопопуляции. Трансбиотические взаимовлияния могут быть:
- отрицательными: взаимодействие гриба и растения-микосимбиотрофа повышает его конкурентоспособность по отношению к растению, не имеющему микоризы
- положительными: фитопаразит, угнетая одно растение, делает другое более конкурентоспособным; скот, поедая на пастбище одни растения и не поедая сорняки, способствует уменьшению обилия первых, и, как следствие, распространению вторых
Влияние фитоценоза на среду
При формировании фитоценоза, благодаря совокупному влиянию входящих в него организмов, первичная абиотическая среда экотопа превращается в фитоценотическую среду (фитосреду)[15], а сам экотоп — в биотоп. Фитоценоз при этом оказывает влияние практически на все абиотические факторы, изменяя их в ту или иную сторону.
Световой режим фитоценоза

Внутри любого фитоценоза световой режим будет отличается от светового режима открытого участка, не занятого растительностью. Такие отличия возникают за счёт того, что свет в фитоценозе определённым образом перераспределяется и происходят следующие процессы:
- отражение части света за пределы фитоценоза
- поглощение части света растениями (в том числе, в процессе фотосинтеза)
- проникновение света внутрь фитоценоза
Вследствие отражения и поглощения света растениями, до уровня почвы доходит лишь незначительная его часть, что особенно хорошо видно в многоярусных лесных фитоценозах: под пологом соснового леса освещённость составляет, в среднем, 25-30 %, дубового — около 3 %, а влажного тропического леса — около 0,2 % от полной освещённости (на открытой поверхности в тех же географических условиях)[11].
Освещённость в фитоценозе неоднородна: она изменяется как в вертикальном, так и в горизонтальном направлении. При движении от верхней границы фитоценоза до уровня почвы освещёноость падает скачкообразно прежде всего вследствие особенностей облиствения (густоты и расположения листьев в пространстве) на каждом ярусе.
В травяных фитоценозах выделяют два типа освещённости: злаковый и двудольный[16]. Злаковый тип характерен для сообществ с преобладанием растений с вертикально ориентированными листьями (злаки, осоки) и характеризуется постепенным снижением освещённости сверху вниз. Двудольный тип — для сообществ с горизонтально ориентированными листьями; характеризуется резкими скачками освещённости и сходен в этом плане с изменением освещённости в лесных сообществах.
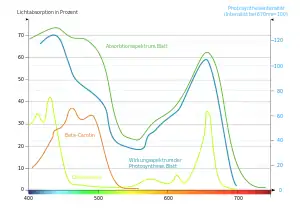
В водных экосистемах в поглощении и отражении света, помимо растений, участвую также вода и взвешенные в ней частицы, вследствие чего на большой глубине существование растений становится невозможным. В прозрачных пресных водоёмах освещённость становится менее 1 % на глубинах более 5-10 метров, вследствие чего высшие растения в таких условиях встречаются на глубинах не более 5, а водоросли — не более 20 метров, однако в прозрачных водах морях и океанах отдельные виды красных водорослей проникают на глубину в несколько сотен метров.
Растения в фитоценозах изменяют и качественный состав спектра, избирательно поглощая и отражая свет с определённой длиной волны. Жёсткое УФ-излучение (λ < 280 нм), вредное для живых структур растительной клетки, практически полностью (до 95-98 %) поглощается эпидермисом листьев и другими покровными тканями. Видимая часть солнечного спектра (фотосинтетически активная радиация) до 70 % поглощается фотосинтетическими пигментами, при этом интенсивнее поглощается сине-фиолетовая и красная части спектра, а зелёная — значительно слабее. ИК-излучение с λ > 7000 нм поглощается до 97 %, а с λ < 2000 нм — очень слабо.
Проницаемость листьев для света также неодинакова и зависит, прежде всего, от толщины и строения листа, а также длины световой волны. Так, листья средней толщины пропускают до 10-20 % света, очень тонкие — до 40 %, а толстые, жесткие, покрытые восковым налётом или опушением листья могут не пропускать свет вовсе. Наибольшей проникающей способностью обладают ИК и зелёный свет, остальные участки спектра проникают через листья существенно меньше.
Световой режим также изменяется в течение суток, в течение года и в зависимости от возрастного состава растений фитоценоза.
Таким образом, фитоценозы изменяют условия освещения экотопа, формируя особый, неоднородный в пространстве световой режим. Специфика этого режима в каждом конкретном фитоценозе определяет набор видов, их распределение и структуру фитоценоза в целом.
Тепловой режим фитоценоза
Участок поверхности Земли, лишённый растительности, получает тепло как непосредственно от Солнца, так и опосредованно — через рассеянный свет неба и обратное излучение нагретой атмосферы. Поступившее тепло частично отражается обратно в атмосферу, частично поглощается и отводится в более глубокие слои Земли и частично излучается нагретой почвой обратно в атмосферу. Все процессы поступления и отведения тепла формируют собой тепловой баланс.
Тепловой баланс меняется в течение суток: днём, в фазу инсоляции, он оказывается положительным, ночью — отрицательным. На тепловой баланс также влияют погодные условия, рельеф местности, время года и географическое положение экотопа.
Фитоценозы существенно изменяют тепловой режим экотопа, так как растения:
- отражают часть солнечного света обратно в атмосферу, снижая поступление тепла в фитоценоз
- поглощают солнечный свет и расходуют его в дальнейшем на физиологические процессы
- выделяют некоторое количество тепла в процессе дыхания
- осуществляют транспирацию и гуттацию
- поглощают часть излучаемого почвой тепла
- снижают испарение с поверхности почвы
- замедляют движение воздушных масс
Также изменение теплового режима экотопа происходит вследствие конденсации и физического испарения влаги с поверхности растений.
Основной энергообмен в фитоценозе осуществляется не у поверхности почвы, как на участке, лишённом растительности, а в верхнем сомкнутом фитоценогоризонте, который сильнее всего нагревается в дневное время и сильнее всего охлаждается в ночное.
В целом, тепловой режим фитоценоза имеет следующие особенности по сравнению с таковым у участка, лишённого растительности:
- максимальные температуры понижаются, а минимальные — повышаются
- суточные и сезонные амплитуды температур оказываются более сглаженными
- среднегодовая температура ниже
Воздушный режим фитоценоза
Влияние фитоценоза на воздушный режим проявляется в изменении скорости движения и состава воздуха. Скорость ветра в фитоценозах падает по направлению сверху вниз и от более открытого участка к менее открытому. Динамика же состава воздуха определяется, главным образом, изменением концентраций кислорода и углекислого газа в процессе фотосинтеза и дыхания. Более значительным колебаниям подвержено содержание CO2: в первой половине дня оно понижается, что связано с интенсификацией фотосинтеза, во второй половине дня — повышается и достигает максимума ночью. Изменение содержания кислорода в воздухе происходит синхронно, но в обратном направлении. Имеется определённая изменчивость содержания CO2 и O2 и по сезонам года: так, в лесах умеренного пояса наименьшая концентрация CO2 наблюдается весной, когда распускаются листья и у растений интенсифицируются процессы жизнедеятельности.
Режим влажности фитоценоза
Структура фитоценоза
В зависимости от специфики исследований в понятии «структуре биоценоза» В. В. Мазингом[17] выделяются три направления, разработанные им для фитоценозов.
1. Структура, как синоним состава (видовая, конституционная). В этом смысле говорят о видовой, популяционной, биоморфологической (состав жизненных форм) и других структурах ценоза, имея в виду лишь одну сторону ценоза — состав в широком смысле. В каждом случае проводится качественный и количественный анализ состава.
2. Структура, как синоним строения (пространственная, или морфоструктура). В любом фитоценозе растения характеризуются определённой приуроченностью к экологическим нишам и занимают определённое пространство. Это относится и к остальным компонентам биогеоценоза. Между частями пространственного расчленения (ярусы, синузии, микрогруппировки и др.) можно достаточно легко и точно провести границы, можно нанести их на план, вычислить площадь, а затем, например, рассчитать ресурсы полезных растений или кормовые ресурсы животных. Только на основе данных по морфоструктуре можно объективно определить точки постановки тех или иных экспериментов. При описании и диагностики сообществ всегда проводится изучение пространственной неоднородности ценозов.
3. Структура, как синоним совокупностей связей между элементами (функциональная). В основе понимания структуры в таком смысле лежит изучение взаимоотношений между видами, в первую очередь изучение прямых связей — биотический коннекс. Это изучение цепей и циклов питания, обеспечивающих круговорот веществ и раскрывающих механизм связей трофических (между животными и растениями) или топических (между растениями — конкуренция за питательные вещества в почве, за свет в надземной сфере, взаимопомощь).
Все три аспекта структуры биологических систем тесно взаимосвязаны на ценотическом уровне: видовой состав, конфигурация и размещение структурных элементов в пространстве являются условием для их функционирования, то есть жизнедеятельности и продуцирования растительной массы, а последнее, в свою очередь, в значительной степени определяет морфологию ценозов. И все указанные аспекты отражает условия среды, в которых формируется биогеоценоз.
Фитоценоз состоит из ряда структурных элементов. Различают горизонтальную и вертикальную структуру фитоценоза. Вертикальная структура представлена ярусами, выделяемыми по визуально определяемым горизонтам концентрации фитомассы. Ярусы состоят из «разновысотных» растений. Примерами ярусов являются 1-й древесный ярус, 2-й древесный ярус, напочвенный покров, мохово-лишайниковый ярус, ярус подлеска и др. Число ярусов может различаться. Эволюция фитоценозов идёт в направлении увеличения числа ярусов, так как это ведёт к ослаблению конкуренции между видами. Поэтому в более древних лесах умеренного пояса Северной Америки число ярусов (8—12) больше, чем в аналогичных более молодых лесах Евразии (4—8).
Горизонтальная структура фитоценоза формируется благодаря наличию пологов деревьев (под которыми формируется среда, несколько отличная от среды в межпологовом пространстве), неоднородностям рельефа (которые обуславливают изменение уровня грунтовых вод, различную экспозицию), видовым особенностям некоторых растений (размножающихся вегетативно и образующих моновидовые «пятна», изменение среды одним видов и реагирование на это другими видами, аллелопатические воздействия на окружающие растения), деятельности животных (например, формирование пятен рудеральной растительности на пороях грызунов).
Закономерно повторяющиеся пятна (мозаики) в фитоценозе, различающиеся составом видов или их количественным соотношением, называются микрогруппировками[18], а такой фитоценоз — мозаичным.
Неоднородность может также носить случайный характер. В этом случае она называется пестротностью.
Динамика фитоценозов
Фитоценозы характеризуются постоянством видового состава, условиями обитания, но там все равно происходят непрерывные изменения. Свойство изменяться называется динамичностью.
Динамические процессы бывают:
А. Обратимые
1. Суточные – связаны с суточной ритмикой жизнедеятельности растений, образующих фитоценоз; выражаются в изменениях активности транспирации, дыхания, фотосинтеза, в суточных движениях цветков и листьев, в ритме открывания и закрывания цветков. Определяются особенностями фитоклимата, создаваемого растительным сообществом.
2. Сезонные – определяются особенностями ритма развития видов, образующих фитоценоз. Эти изменения позволяют существовать совместно большему числу видов растений, чем в том случае, если бы они развивались одновременно. Н-р, весной – ранние эфемероиды, летом – позднелетние травы, кустарники, деревья.
3. Флуктуации — это разногодичные изменения, связанные с неодинаковыми условиями существования растений в разные годы. Состав не изменяется, может меняться численность и возрастной состав популяции.
Б. Необратимые(сукцессии, эволюция сообществ, нарушения сообществ).
Сукцессии — это постепенные изменения фитоценозов необратимые и направленные, вызванные внутренними или внешними причинами, по отношению к фитоценозам, причинам. Выделяются первичные и вторичные сукцессии. Первичные сукцессии начинаются на безжизненных субстратах (скалах, обрывах, наносах рек, сыпучих песках), вторичные же начинаются на субстратах, на которых растительность была, но нарушена (восстановление после лесного пожара).
Классификация фитоценозов
При классификации фитоценозов сходные сообщества объединяют в группы — классификационные единицы.
Низшая единица классификации — ассоциация (совокупность однородных фитоценозов, имеющих более или менее одинаковый внешний облик, сходный флористический состав и одни и те же доминирующие виды по ярусам). Названия ассоциаций даются перечислением русских названий доминирующих растений каждого яруса фитоценоза начиная с самого верхнего яруса (сосна обыкновенная + ель европейская — брусника + черника — мох плеуроциум) или латинских родовых и видовых названий доминантов (Pinus sylvestris + Picea abies — Vaccinium vitis-idaea + Vaccinium myrtillus — Pleurozium schreberi) с прибавлением к основе суффиксов лат. -etum, -osum, -estosum: Piceetum oxalidosum (от Picea и Oxalis) — ельник кисличный.
Формация — это совокупность ассоциаций, у которых в верхнем ярусе доминирует один и тот же вид растения (например, сосняки, дубравы и пр.)
Ординацией называют построение рядов фитоценозов по постепенному изменению какого-либо фактора окружающей среды в определённом направлении. Так, можно провести ординацию по фактору увлажнения почвы. В этом случае получится ряд сообществ, где каждое займет соответствующее место в зависимости от условий увлажнения, в которых оно развивается, причём крайнее из них будет соответствовать наиболее влажным почвам, а противоположное — наиболее сухим.
Эдафо-фитоценотические ряды типов еловых лесов (по В. Н. Сукачеву, 1972):
1) Ельники-зеленомошники. Встречаются в условиях расчленённого рельефа, на более или менее богатых суглинках, а также глинах и супесях. К этой группе относят ельники кисличные, черничные и брусничные.
2) Ельники-долгомошники (в моховом ярусе господствует кукушкин лён). Встречаются в условиях более или менее равнинного рельефа на слабо дренированных почвах с признаками заболачивания (состав почв тот же, что и в ельниках первой группы).
3) Ельники сфагновые. Встречаются в условиях равнинного рельефа, нередко в котловинных понижениях, на недренированных заболоченных почвах. Моховой покров состоит в основном из сфагновых мхов.
4) Ельники травяные. Располагаются по дну долин небольших речек и ручьёв в виде узких полос на богатых сырых почвах с проточным увлажнением. Хорошо развит травяной покров.
5) Ельники сложные. Приурочены к богатым, хорошо дренированным почвам, часто с близким залеганием известняков. В этих лесах хорошо развиты подлесок и травяной покров, присутствуют широколиственные породы — дуб, клён, липа.
Эти пять групп ассоциаций образуют экологические ряды, отражающие изменения еловых лесов в зависимости от увлажнения и богатства почвы. Ряд А (от кисличного к брусничному ельнику) — увеличение сухости почвы и некоторое обеднение почвы. Ряд В (от кисличного к сфагновому ельнику) — увеличение увлажнения и ухудшения аэрации почвы. Ряд С (от кисличного к сложному ельнику) — увеличение богатства и аэрации почвы. Ряд Д (от кисличного к приручьёвому ельнику) — увеличение проточного увлажнения. Ряд Е (от кисличного к сфагновому ельнику) — увеличение застойного увлажнения и уменьшение аэрации почвы.
См. также
Примечания
- Баркман Н. Современные представления о непрерывности и дискретности растительного покрова и природе растительных сообществ в фитосоциологической школе Браун-Бланке. — Ботанический журнал, 1989, т. 74 № 11.
- Раменский Л. Г. О сравнительном методе экологического изучения растительных сообществ // Дневник съезда русских естествоиспытателей и врачей — СПб., 1910.
- Greason H. A. The structure and development of the plant assotiation // Bul. Torrey Bot. Club — 1917.
- Negri G. Le unita ecologiche fundamentali in fitogeografia — Roy. Acad Sc. Torino, 1914.
- Lenoble F. A propos des associations vegetales // Bull. Soc. Bot. France — 1926.
- Александрова В. Д. О единстве непрерывности и дискретности в растительном покрове // Философские проблемы современной биологии. — М.-Л.: Наука, 1966.
- Александрова В. Д. Классификация растительности. — Л.: Наука, 1969.
- Сукачёв В. Н. Дендрология с основами лесной геоботаники. — Л.: Гослестехиздат, 1938.
- Шенников А. П. Введение в геоботанику. — Л.: Изд-во ЛГУ, 1964.
- Weaver J. E., Clements F. E. Plant ecology. — N.-Y.-London, 1938.
- Прокопьев Е. П. Введение в геоботанику. Учебное пособие. — Томск: Изд-во ТГУ, 1997. — 284 с.
- Работнов Т. А. Фитоценология. — М.: Изд-во МГУ, 1978.
- Работнов Т. А. История фитоценологии. — М.: Аргус, 1995.
- Тихнова Г. Н. Фикусы-удушители с острова Борнео (недоступная ссылка)
- Фитоценотическая среда//Экологический энциклопедический словарь. — Кишинев: Главная редакция Молдавской советской энциклопедии. И. И. Дедю. 1989.
- Лархер В. Экология растений. — М., Мир, 1978
- Мазинг В. В. Что такое структура биогеоценоза // Проблемы биогеоценологии. — М.: Наука, 1973. — С. 148–156.
- Ярошенко П. Д. Геоботаника: Пособие для студентов педвузов. — М.: Просвещение, 1969. — 200 с.
Литература
- Любарский Е. Л. Ценопопуляция и фитоценоз. — Казань, 1976.
