Клеточное дыхание
Клеточное, или тканевое дыхание — совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, липидов и аминокислот до углекислого газа и воды, а также образование энергии. Высвобожденная энергия запасается в химических связях макроэргических соединений (АТФ, которых в результате процесса образуется 30 (32) и др.) и может быть использована по мере необходимости. Входит в группу процессов катаболизма. О физиологических процессах транспортировки к клеткам многоклеточных организмов кислорода и удалению от них углекислого газа см. статью Дыхание.
Использование различных начальных субстратов
В качестве исходных субстратов дыхания могут выступать различные вещества, преобразуемые в ходе специфических метаболических процессов в Ацетил-КоА с высвобождением ряда побочных продуктов. Восстановление НАД (НАДФ) и образование АТФ может происходить уже на этом этапе, однако большая их часть образуется в цикле трикарбоновых кислот при переработке Ацетил-КоА.
Гликолиз
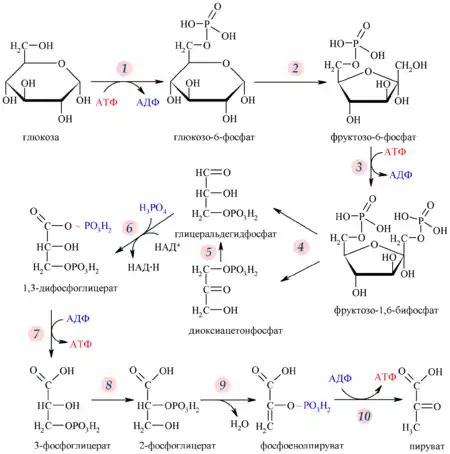
Гликолиз — путь ферментативного расщепления глюкозы — является общим практически для всех живых организмов процессом. У аэробов он предшествует собственно клеточному дыханию, у анаэробов завершается брожением. Сам по себе гликолиз является полностью анаэробным процессом и для осуществления не требует присутствия кислорода.
Первый его этап протекает с высвобождением 2 молекул АТФ и включает в себя расщепление молекулы глюкозы на 2 молекулы глицеральдегид-3-фосфата. На втором этапе происходит НАД-зависимое окисление глицеральдегид-3-фосфата, сопровождающееся субстратным фосфорилированием, то есть присоединением к молекуле остатка фосфорной кислоты и формированием в ней макроэргической связи, после которого остаток переносится на АДФ с образованием АТФ.
Таким образом, уравнение гликолиза имеет следующий вид:
Сократив АТФ и АДФ из левой и правой частей уравнения реакции, получим:
Окислительное декарбоксилирование пирувата
Образовавшаяся в ходе гликолиза пировиноградная кислота (пируват) под действием пируватдегидрогеназного комплекса (сложная структура из 3 различных ферментов и более 60 субъединиц) распадается на углекислый газ и ацетальдегид, который вместе с Коферментом А образует Ацетил-КоА. Реакция сопровождается восстановлением НАД до НАД∙Н.
У эукариот процесс протекает в матриксе митохондрий.
β-окисление жирных кислот
Деградация жирных кислот (у некоторых организмов также алканов) происходит у эукариот в матриксе митохондрий. Суть этого процесса заключается в следующем. На первой стадии к жирной кислоте присоединяется кофермент А с образованием ацил-KoA. Он дегидрируется с последовательным переносом восстановительных эквивалентов на убихинон дыхательной ЭТЦ. На второй стадии происходит гидратирование по двойной связи С=С, после чего на третьей стадии происходит окисление полученной гидроксильной группы. В ходе этой реакции восстанавливается НАД.
Наконец, на четвёртой стадии образовавшаяся β-кетокислота расщепляется β-кетотиолазой в присутствии кофермента А на ацетил-КоА и новый ацил-КоА, в которой углеродная цепь на 2 атома короче. Цикл β-окисления повторяется до тех пор, пока вся жирная кислота не будет переработана в ацетил-КоА.
Цикл трикарбоновых кислот
Ацетил-КоА под действием цитратсинтазы передаёт ацетильную группу оксалоацетату с образованием лимонной кислоты, которая поступает в цикл трикарбоновых кислот (цикл Кребса). В ходе одного оборота цикла лимонная кислота несколько раз дегидрируется и дважды декарбоксилируется с регенерацией оксалоацетата и образованием одной молекулы ГТФ (способом субстратного фосфорилирования), трёх НАДН и ФАДН2.
Суммарное уравнение реакций:
У эукариот ферменты цикла находятся в свободном состоянии в матриксе митохондрий, только сукцинатдегидрогеназа встроена во внутреннюю митохондриальную мембрану.
Окислительное фосфорилирование
Основное количество молекул АТФ вырабатывается по способу окислительного фосфорилирования на последней стадии клеточного дыхания: в электронтранспортной цепи. Здесь происходит окисление НАД∙Н и ФАДН2, восстановленных в процессах гликолиза, β-окисления, цикла Кребса и т.д. Энергия, выделяющаяся в ходе этих реакций, благодаря цепи переносчиков электронов, локализованной во внутренней мембране митохондрий (у прокариот — в цитоплазматической мембране), трансформируется в трансмембранный протонный потенциал. Фермент АТФ-синтаза использует этот градиент для синтеза АТФ, преобразуя его энергию в энергию химических связей. Подсчитано, что молекула НАД∙Н может дать в ходе этого процесса 2.5 молекулы АТФ, ФАДН2 — 1.5 молекулы.
Конечным акцептором электрона в дыхательной цепи аэробов является кислород.
Анаэробное дыхание
Если в электронтранспортной цепи вместо кислорода используется другой конечный акцептор (трёхвалентное железо, нитрат- или сульфат-анион), дыхание называется анаэробным. Анаэробное дыхание свойственно в основном бактериям, которые благодаря этому играют важную роль в биогеохимическом цикле серы, азота и железа. Денитрификация — один из типов анаэробного дыхания — является одним из источников парниковых газов, железобактерии принимают участие в образовании железомарганцевых конкреций. Среди эукариот анаэробное дыхание встречается у некоторых грибов, морских донных беспозвоночных, паразитических червей[1] и протистов — например, фораминифер[2].
Общее уравнение дыхания, баланс АТФ
| Стадия | Выход кофермента | Выход АТФ (ГТФ) | Способ получения АТФ |
|---|---|---|---|
| Первая фаза гликолиза | −2 | Фосфорилирование глюкозы и фруктозо-6-фосфата с использованием 2 АТФ из цитоплазмы. | |
| Вторая фаза гликолиза | 4 | Субстратное фосфорилирование | |
| 2 НАДН | 3 (5) | Окислительное фосфорилирование. Только 2 АТФ образуется из НАДН в электронтранспортной цепи, поскольку кофермент образуется в цитоплазме и должен быть транспортирован в митохондрии. При использовании малат-аспартатного челнока для транспорта в митохондрии из НАДН образуется 3 моль АТФ. При использовании же глицерофосфатного челнока образуется 2 моль АТФ. | |
| Декарбоксилирование пирувата | 2 НАДН | 5 | Окислительное фосфорилирование |
| Цикл Кребса | 2 | Субстратное фосфорилирование | |
| 6 НАДН | 15 | Окислительное фосфорилирование | |
| 2 ФАДН2 | 3 | Окислительное фосфорилирование | |
| Общий выход | 30 (32) АТФ[3] | При полном окислении глюкозы до углекислого газа и окислении всех образующихся коферментов. | |
См. также
Примечания
- Tielens A.G.M., Rotte C., van Hellemond J.J., Martin W. Mitochondria as we don't know them (Trends in Biochem.Sci.,2002,27,11,564-572
- Если нет кислорода, можно дышать нитратами
- David L. Nelson, Michael M. Cox. Lehninger Principles of Biochemistry. — 4. — W. H. Freeman, 2004. — 1100 с.