Тоди
То́ди[1] (лат. Todus) — единственный род птиц семейства тодиевых (Todidae), в который включают пять видов, обычно эндемичных для какого-то одного из Больших Антильских островов. Они могут жить на любых высотах от почти 50 метров ниже уровня моря до 3000 метров над уровнем моря, предпочитают места с переплетёнными ветвями и лианами, что связано с редкими перелётами тоди на расстояния больше 1—2 метров. Излюбленным местом обитания являются тенистые кофейные плантации, на которых кофейные кусты растут в тени высоких лиственных деревьев.
| Тоди | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Ямайский тоди | ||||||||||||
| Научная классификация | ||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Класс: Подкласс: Инфракласс: Клада: Отряд: Семейство: Тодиевые (Todidae Vigors, 1825) Род: Тоди |
||||||||||||
| Международное научное название | ||||||||||||
| Todus Brisson, 1760 | ||||||||||||
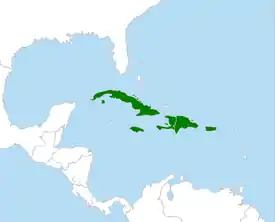
| Ареал тодиевых | ||||||||||||
 |
||||||||||||
| ||||||||||||
Небольшие птицы общей длиной 10—11 сантиметров и массой 4,3—10,2 грамма имеют ярко-зелёное оперение в верхней части тела и беловатую грудку с розовыми боками, ярко-красным горловым пятном и жёлтым подхвостьем. Для них характерен длинный уплощённый клюв с тонкими зазубринами по краю верхней части, которые помогают разламывать твёрдых насекомых — основной корм тоди. Для защиты территории могут издавать крыльями звук, похожий на трещотку, когда воздух быстро проходит через первостепенные маховые перья. Гнездятся в основном в норах, которые копают клювом и лапами оба партнёра, поэтому тоди имеют частично сросшиеся фронтальные пальцы.
Общая характеристика
Описание
Тоди — птицы округлой формы тела с крупной головой и относительно длинным клювом[2]. Их общая длина составляет в среднем 10—11 см (по данным 1914 года — 87—110 мм[3]), масса — 4,3—10,2 г[4]. Тоди имеют блестящее ярко-зелёное оперение верхней части тела, крыльев и хвоста; беловатое оперение нижней части тела с ярко-красным горловым пятном, которое отсутствует у молодых особей; кроющие перья подхвостья жёлтые, бока — розовые, интенсивность цвета зависит от вида тоди[2].
Крылья у тоди короткие и округлые. Это связано с тем, что птицы ведут оседлый образ жизни и не перелетают на большие расстояния[5]. Самыми длинными обычно являются седьмое и восьмое маховые перья; девятое перо короче пятого, а иногда и шестого пера; длина десятого — внешнего — махового пера составляет примерно три четверти от максимальной[3]. При этом общая длина крыла тесно связана со средней продолжительностью перелётов во время охоты; в частности, ширококлювый тоди, длина крыла которого составляет 50,3 мм, имеет наибольшее среднее расстояние полёта — 2,2 метра. Ему же принадлежит и максимальное зарегистрированное расстояние полёта — 40 метров. Остальные виды тоди обладают более короткими крыльями, длиной 42,8—46 мм, а среднее и максимальное расстояние полёта для них составляют 0,8—1,5 метров и 7—26 метров соответственно[5]. Хвост птиц имеет широкое закруглённое окончание, его длина составляет от двух третей до трёх четвертей длины крыла[3].
Особых изменений в окраске оперения в течение года у взрослых птиц не наблюдается. Исследования линьки, по-видимому, проводились только для ямайского тоди и показали, что птицы меняют перья после сезона размножения, с начала августа. При этом все маховые перья меняются к концу октября, а смена остальных перьев продолжается ещё два месяца[5].
Клюв у тоди длинный, прямой и слегка приплюснутый, его длина вдвое меньше длины крыла. У большинства видов клюв широкий, с закруглённым концом, его ширина в районе ноздрей примерно в два раза больше толщины[3]. Исключение составляет узкоклювый тоди. Нижняя часть клюва обычно имеет красный цвет[2], а верхняя — чёрный[5]. Верхняя часть клюва у тоди неглубокая для того, чтобы снимать насекомых с листьев[5], по её краю расположены зазубрины, с помощью которых птицы разламывают твёрдых насекомых[2]. Длина клюва от ноздрей колеблется в диапазоне 12,3—14,3 мм, ширина в районе ноздрей составляет 3,9—5,4 мм, а высота — 2,1—3,7 мм. Самым маленьким клювом обладает кубинский тоди, самым большим — ширококлювый[5].
Лапы примерно той же длины, что и клюв, длиннее среднего пальца с когтем. Внешний палец слегка короче среднего, внутренний — намного короче; большой палец примерно такой же длины, что и внутренний. Все фронтальные пальцы частично объединены между собой (до предпоследнего сгиба)[3] — характерная черта для всех ракшеобразных, так как это необходимо для копания нор. Когти большие, острые, сильно закруглённые[3]. Кроме того, лапы тоди приспособлены для сидения на мелких веточках[5].
Половой диморфизм в целом отсутствует[4], однако наблюдается корреляция между полом и цветом радужки. Более всего это выражено у пуэрто-риканского тоди, в то время как у птиц с острова Гаити цвет радужки связан с определением вида, а у кубинского тоди — с возрастом птиц[5].
Вокализация
Тоди исполняют простые песни с заметным носовым звучанием, сонограммы их вокализаций являются одними из самых простых среди всей авифауны. Исключение составляет кубинский тоди, чья песня состоит из четырёх слогов. Помимо этого, тоди могут издавать горловые звуки. Во время дождя вокализация, как и общая активность, снижается. Самыми тихими считают ямайских тоди[6].
Все тоди обычно летают бесшумно, но для защиты территории могут издавать характерный звук крыльями, когда воздух быстро проходит через первостепенные маховые перья[2]. Ранее считалось, что этот звук связан с тем фактом, что перья слегка ослаблены, однако при тщательном изучении оказалось, что эти перья не обладают изменённой жёсткостью или шириной. Довольно громкий звук вместе с тем трудно поддаётся записи и сильно зависит от погодных условий: в солнечные дни он слышен сильнее, чем в пасмурные или дождливые. Этот звук, напоминающий трещотку, похож на звук, который птицы издают горлом. Чаще всего его можно услышать в сезон размножения во время территориальных споров или брачных игр[6].
Впервые звук, издаваемый крыльями, был описан в 1943 году американским герпетологом Томасом Барбуром применительно к кубинскому виду[6]:
Тоди — это крошечные живые существа, гораздо меньшие, чем английский воробей [Passer domesticus],… и каждый раз, когда они перелетают, их крылья трещат при каждом взлёте, как у гигантского кузнечика.
Оригинальный текст (англ.)[показатьскрыть]The todies are tiny verdigris busybodies, far smaller than an English Sparrow [Passer domesticus]… and as they make each foray their wings snap like those of a giant grasshopper at each takeoff.
Распространение
Тоди обитают на Антильских островах. Ареалы трёх видов не пересекаются: кубинский тоди обитает на острове Куба (а также соседнем острове Хувентуд), пуэрто-риканский — на острове Пуэрто-Рико, ямайский — на острове Ямайка, и только узкоклювый и ширококлювый тоди обитают на острове Гаити, в западной и восточной частях острова, соответственно[2].
Тоди могут жить на любых высотах от почти 50 метров ниже уровня моря до 3000 метров над уровнем моря. Они предпочитают леса с переплетёнными ветвями и лианами. Исследования на Кубе показали, что тоди не привязаны к конкретному типу леса. Могут обитать в лесах на известняковых карстах, в горных тропических лесах, в сосновых лесах (Pinus occidentalis, Pinus caribaea), прибрежных лесных массивах и манграх. Встречаются на затенённых кофейных плантациях, бобовых деревьях и кактусах Cephalocereus. Крайне редко встречаются в редколесье, где расстояние между деревьями превышает 3 метра и нет кустарникового покрова. Это связано с тем, что тоди редко перелетают на расстояния больше 1—2 метров. Также редки в пальмовой растительности, где мала плотность насекомых. Кроме того, на среду обитания тоди влияет наличие овражков и уступов, подходящих для копания нор и гнездования[7].
Предпочитают тенистые участки[4]. Большинство видов обитают во влажных лесах, в том числе в гористой местности. Исключение составляют ширококлювый и пуэрто-риканский тоди[2]. Узкоклювый тоди, вероятно, единственный вид, который имеет явные предпочтения в среде обитания. Его можно встретить в сосновых лесах или в высокогорных тропических лесах, где подлесок включает поднимающийся по скалам бамбук, папоротники, эпифиты, печёночные мхи и орхидеи. Ширококлювый тоди, напротив, встречается повсеместно. Эти два вида делят один остров, их ареал пересекается. В Сьерра-де-Баоруко оба вида встречаются в смешанных стаях в вертикальном диапазоне не менее 1200 м, что приводит к заметным отклонениям в их поведении и тактике охоты. Ямайский тоди избегает соснового леса и часто встречается в районах с перемежающимися лесными участками и открытыми пастбищами. В погоне за насекомыми он может залететь в дом[7].
Плотность популяции тоди на территории зависит от таких факторов, как обилие насекомых, густота растительного покрова и возвышенность. В целом площадь территории значительно уменьшается с увеличением высоты. Наибольшая плотность тоди достигается на затенённых кофейных плантациях и в сухих низкогорных лесах. В Лукильо в Пуэрто-Рико на высоте 130—1165 метров размер территории пары составляет в среднем 0,7 га[7].
Тоди присоединяются к смешанным стаям в основном весной и осенью. В это время в таких стаях встречается много мигрирующих лесных певунов (Parulidae), а сами тоди не имеют пар. Смешанные стаи мелких птиц в Карибском бассейне сильно отличаются от таковых в Центральной и Южной Америке меньшим разнообразием. В Пуэрто-Рико тоди обычно присоединяются к стаям с Nesospingus speculiferus, банановыми певунами (Coereba flaveola), дроздами (Turdus), виреонами (Vireo), объединяющим в среднем шесть видов. На Гаити, где видовое разнообразие больше, стаи в среднем включают 11 видов, наиболее частым из которых является Phaenicophilus palmarum[8].
Питание
Рацион
Тоди являются насекомоядными птицами. Новозеландская естествоиспытательница Анджела Кеплер в своих исследованиях, опубликованных в 1977 году, определила 49 семейств насекомых, которых они употребляют в пищу[9]. Основу рациона тоди составляют кузнечики и сверчки (Orthoptera), жуки (Coleoptera), полужесткокрылые (Hemiptera, Homoptera), мотыльки и бабочки (Lepidoptera), мухи (Diptera), тараканы (Dictyoptera), сетчатокрылые (Neuroptera), пчёлы, осы и муравьи (Hymenoptera). Помимо этого, тоди употребляют в пищу пауков, нематод и многоножек, а также яйца насекомых, небольших ящериц и животных[10]. Кроме того, они могут изредка поедать мягкие плоды[2] брунеллиевых (Brunelliaceae), маревых (Chenopodiaceae), клузиевых (Guttiferae), тутовых (Moraceae), маковых (Papaveraceae), розовых (Rosaceae) и мареновых (Rubiaceae). Таким образом, рацион тоди мало пересекается с рационом других позвоночных с Антильских островов и более всего схож с питанием лесных тиранчиков (Hemitriccus), обитающих на континенте. Тоди значительно менее активны, чем лесные певуны, их броски составляют менее 1,5 метров в длину. Наиболее активным представителем является узкоклювый тоди, наименее активным — ширококлювый[10].
Тоди довольно прожорливы: в тропическом лесу Эль-Верде в Пуэрто-Рико семья тоди в течение пяти с половиной месяцев (это средний период, в течение которого пара защищает свою территорию) употребила в пищу приблизительно 1,8 млн насекомых на участке размером 0,7 га. Вместе с тем их метаболизм не является уникальным, а сопоставим с таковым других птиц очень малого размера, в частности, колибри-герцога (Eugenes fulgens) и желтоголового королька (Regulus regulus). Ранее считалось, что скорость обмена веществ тоди очень высока. Такие выводы основывались на наблюдении птицы в неволе, которая ежедневно потребляла примерно 40 % массы своего тела[2], а также обширного исследования, включавшего несколько тысяч наблюдений за питанием всех видов тоди, которое показало, что они съедают в среднем 1,9 насекомого в минуту в засушливых кустарниках и 1,1 — в тропическом лесу[10].
Особенности охоты
.jpg.webp)
Тоди используют несколько техник охоты на насекомых. При «underleaf-sally» клюв и глаза направлены вверх, птицы сканируют нижнюю часть листьев и веток над головой, на добычу бросаются под небольшим углом вверх, продолжая полёт по той же траектории и не возвращаясь. Другая техника, «sally-pounce», связана со сканированием верхней стороны листьев, когда птицы бросаются на добычу вниз, и чаще используется в засушливых кустарниках. Помимо этого, птицы могут зависать в воздухе («sally-hover»), что напоминает охотничьи приёмы мухоловок, а также использовать другие методы охоты, характерные для насекомоядных, как то: щёлканье, подпрыгивание, движения в сторону. Тоди ловят добычу на листьях, папоротниках, стволах, ветках, соцветиях и плодах, реже подбирают с земли или захватывают водных насекомых, подёнок (Ephemeroptera) и стрекоз (Zygoptera)[10].
Тоди предпочитают охотиться в нижних и средних слоях леса, при этом сидят на любых поверхностях, достаточно крепких, чтобы удержать их вес: веточках и ветвях, лозах, черешках, папоротниках и цветах. Диаметр веток может составлять от 1 до 60 мм (5 мм — средний диаметр для пуэрто-риканского тоди, 11 мм — для ширококлювого[5])[10]. Средняя высота кормления составляет 2,3 метра в кустарниковых зарослях и 4,9 м в тропическом лесу. Ширококлювый тоди питается выше, чем другие виды, и поднимается ещё выше в тех областях, где встречается наряду с узкоклювым видом. Это разделение отражается не только в высоте добычи, но и в продолжительности и частоте полёта: ширококлювый вид практикует более длинные перелёты и захватывает более крупную добычу[10].
Обмен веществ
Тоди, как и все маленькие птицы, сталкиваются с проблемой экономии энергии и тепла. Они обладают высокой скоростью обмена веществ, быстрым теплообменом и ограниченной способностью к накоплению запасов энергии за счёт хранения жира. При этом во время сильного дождя у них ограничено время кормления из-за рисков намочить оперение, особенно на больших высотах. Тоди — одни из самых маленьких активно кормящихся птиц — разработали эффективные способы сохранения энергии. В отличие от остальных ракшеобразных, которые предпочитают проводить ночное время в группах, прижавшись друг к другу, тоди больше полагаются на физиологические адаптации, такие как низкие температуры обмена веществ и оцепенение. В частности, температура тела пуэрто-риканского тоди составляет 36,7 °C (температура тела других ракшеобразных — 40 °C), что позволяет снизить ожидаемые затраты энергии на 33 %[8].
Размножение
Ухаживание

Тоди сезонно моногамны, то есть не меняют пару в течение одного сезона[4]. Ухаживания включают преследования друг друга на прямых участках и параболических дугах среди низкой листвы, во время которых птицы покрывают расстояния до 10 метров. Такие игры включают множественные контакты тела и характерные звуки от махов крыльями. Своим поведением в этот сезон тоди напоминают манакинов, которые совершают похожие манёвры, но на открытых участках, а не в густой растительности. Сидя на ветках, птицы ведут себя взволнованно, часто подпрыгивают и распушают оперение[11].
Ещё одной особенностью ухаживания, общей для всех видов, кроме пуэрто-риканского, является демонстрация боков («flank display»), когда розовые боковые перья распушены с различной степенью интенсивности. Во время полёта яркие розовые перья почти соприкасаются за спиной, а крылья едва заметны. Самые характерные проявления этого способа ухаживания принадлежат ширококлювому тоди, за которым следуют по мере уменьшения количества розовых перьев на боках кубинский, узкоклювый и ямайский. Пуэрто-риканский тоди, у которого бока жёлтые, крайне редко распушает своё оперение подобным образом и никогда не щёлкает крыльями за спиной[11].
Тоди редко обмениваются добычей во время ухаживания. После удачной охоты они могут приземлиться или зависнуть перед партнёром и показать ему пойманное насекомое, повторяя такой манёвр 2—3 раза. Совместное питание усиливает связь пары во время инкубационного периода и ухаживания за птенцами. Самка приглашает самца к спариванию, хлопая крыльями, затем расправляя их и поднимая свой короткий хвост[11].
Пары на более мелких территориях защищают их чаще и активнее, чем те, которые занимают более крупные[7].
Гнездование

Первое описание особенностей гнездования тоди было дано в Пуэрто-Рико в 1846 году немецким ботаником Карлом Морицем[11]:
В тенистых деревьях время от времени можно увидеть очаровательных зелёных птичек, хрипло шумящих… Местные жители полагают, что они гнездятся в норах под землёй.
Оригинальный текст (англ.)[показатьскрыть]In shady trees is seen once in a while the lovely green San Pedrito, rattling hoarsely… The locals believe that it nests in holes in the earth.
Копание нор характерно также для момотовых и зимородковых[2], при этом длина туннеля у тоди является одной из самых коротких и составляет от 12 до 60 см, размеры отверстия — 3,4—4,4 см[11], размеры выводковой камеры — 7,5 × 12,5 см[2]. Тоди используют невысокие откосы и овраги с естественным уклоном. Обычно их высота составляет менее 1,5 метров. Чаще всего птицы используют откосы с умеренной растительностью, которая обеспечивает стабилизацию почвы и даёт частичную защиту от мангустов, но могут выкапывать норы и на голых склонах. Обильная растительность препятствует созданию туннеля, так как тоди иногда приходится отрывать несколько сантиметров толстых мхов, чтобы добраться до подходящего грунта. Туннели, сделанные тоди, обычно горизонтальны и заканчиваются выводковой камерой без дополнительного наполнения. Иногда тоди делают норы в мёртвой древесине или используют старые дупла, сделанные дятлами[11]. Роют норы оба партнёра[2].
Тоди начинают примерно в три раза больше нор, чем используют, из-за неподходящей почвы, включая недостаточный растительный покров, наличия препятствий в виде камней, корней или быстрорастущих папоротников, а также приливов после сильных дождей. Они не возвращаются в затопленные гнёзда. При этом, поскольку тоди ведут преимущественно оседлый образ жизни, они могут использовать норы, вырытые в предыдущие годы, но при этом ежегодно вырывают и свежие туннели[11].
В целом тоди занимаются выкапыванием норы с сентября по июнь, при этом основная активность приходится на период с февраля по май. Тоди, обитающие в тропических лесах, обычно строят гнёзда и откладывают яйца в сухой сезон, а тоди, обитающие в засушливых кустарниках, — наоборот, в сезон дождей. Каждая нора копается в течение восьми недель. В начале копания птицы пользуются клювом как долотом, посещая нору до 60 раз в час, а затем возвращаясь на любимую ветку. На следующей стадии тоди копают нору лапами. При этом по утрам в течение часа птица роет нору 15 минут, а затем 45 минут занимается кормлением. Рытьё полностью прекращается днём и во время дождей или облачности[11].
Пара много общается между собой во время рытья норы. Самец и самка работают попеременно. Также оба пола участвуют в защите территории от нарушителей. Аккуратные норы привлекают внимание многочисленных мелких животных, среди которых листовые лягушки (Eleutherodactylus), анолисы (Anolis), Tityrus, Solenopsis, Amphicausta, Cyrtophilis. В качестве стратегии против избегающих света хищников тоди строят свои туннели на солнечной стороне[11]. После того как тоди покидают нору, её обычно занимают другие животные[2].
Птенцы
Яйца тоди откладывают раз в год, в кладке 1—4 крупных яйца белого цвета. Из-за большого оранжево-красного желтка яйца окрашиваются в розовый цвет. Размер яиц тоди схож с соответствующими размерами у крапивников (Troglodytes) и немного больше яиц у колибри. Яйцо весит 1,4 грамма, что составляет 26 % массы взрослой птицы. В случае уничтожения яиц птицы могут попытаться отложить вторую кладку[11]. Основным естественным хищником для тоди является малый мангуст (Urva auropunctata), который лакомится их яйцами. По некоторым данным, до 81 % разорённых гнёзд в Лукильо пострадали именно от мангустов[12].
Предположительно, инкубационный период длится 21—22 дня. Высиживают птенцов оба партнёра, но они постоянно меняются, так как тоди должны кормиться чаще, чем более крупные птицы. Поскольку взрослые птицы редко пересекаются в гнезде, они предпочитают залетать в него и покидать его бесшумно. Изредка можно услышать короткие звуковые сигналы или звуки от крыльев. Вылупление происходит главным образом во второй половине дня и занимает менее одного часа, при этом инкубация начинается с первого яйца[11]. В первую неделю горло птенцов обычно бледно-серое, а длина клюва примерно в два раза меньше, чем у взрослой птицы. Полные размеры достигаются в течение по меньшей мере трёх недель[5].
Птенцы кормятся в основном насекомыми[10] и издают писк на высокой частоте в ожидании еды[6]. Родители показывают удивительное усердие в доставке пищи. Известно о паре, которая доставляла своему выводку 420 насекомых в день. При этом размер насекомых зависит от размеров птенцов, а основную диету составляют Homoptera, Coleoptera, Lepidoptera, Orthoptera и Hemiptera[11].
За два-три дня до вылета птенцов из гнезда взрослые птицы по несколько часов подряд парят около выхода из него, издавая звуковые сигналы, подпрыгивают и действуют взволнованно. В ответ птенцы тоже громко шумят. Птенцы никогда не возвращаются к гнезду, хотя в течение первых пяти недель не отлетают от него далее, чем на 25 метров. Родители ещё кормят птенцов в течение трёх недель после вылета из гнезда, главным образом в первую неделю, после чего они выкладывают пищу на небольшом расстоянии от птенца, заставляя последнего двигаться, чтобы получить еду. Из-за этих особенностей взросление птенцов тоди занимает довольно много времени, полное формирование происходит примерно через 42 дня, а шум крыльями появляется после четырёх месяцев[11].
Мало известно о максимальной продолжительности жизни тоди. Есть сведения о птицах, проживших семь-восемь лет, но неизвестно, насколько этот показатель является типичным для семейства[11].
Тоди и человек
Свои местные названия представители семейства получили из-за особенностей гнездования — barrancolí, barranquero или barrancolino от испанского слова barranco (овраг) или perroquet de terre, что в переводе с французского означает «земельный попугай»; или вокализации — pichui, chicorette, papagayo из-за громкого шума, pedorrera на Кубе из-за звука, издаваемого крыльями. Малые размеры птицы послужили причиной для названия medio peso в Пуэрто-Рико или colibri на Гаити, из-за чего существует опасность перепутать тоди с колибри. Англичане на Ямайке назвали тоди robin redbreast, основываясь на примере зарянки (Erithacus rubecula) и ярко-красном горловом пятне[13].

В настоящее время все виды тоди находятся под наименьшей угрозой, однако с 1988 по 2000 год узкоклювого тоди относили к видам, близким к уязвимому положению[12][14].
Ранее человеческая деятельность шла на пользу тоди. Птицы могли копать норы в срытых для дорог уступах или дренажных каналах, предпочитали тенистые кофейные плантации. Выращивание кофе очень популярно на Антильских островах. Ранее для этого использовали метод Cafetales, когда кофейные кусты росли в тени высоких лиственных деревьев, сохраняя при этом естественный ландшафт с возможностями для копания нор. В настоящее время кофе садят на солнце, а плантации ровняют бульдозером. Очевидно, что на популяцию тоди также оказывает влияние количество насекомых. Больше всего их на низких высотах, которые были ранее всего культивированы человеком. Современный туристический бизнес заставляет использовать в больших количествах различные спреи от насекомых, в частности, малатион на Кайо-Коко, что также сказывается на популяции насекомоядных тоди. Фактор глобального потепления увеличивает количество дождей в Карибском бассейне, вследствие чего тоди имеют проблемы с поиском пропитания, а их норы могут оказаться затоплены[12].
Яйца тоди служат источником протеина для местных детей, которые могут легко добраться до нор, расположенных вдоль дорог[12].
Тоди совсем не живут в садах, известно только несколько подобных случаев: на Ямайке однажды тоди отложили яйца в деревянном цветочном горшке, нору к которому прорыли сами, а на острове Хувентуд близ Кубы тоди обитал в грейпфрутовом саду. Тоди практически не размножаются в неволе, однако человек может поддерживать тоди в естественной среде, создавая искусственные невысокие откосы, особенно на кофейных плантациях[12].
Эволюция
Происхождение семейства
Самыми старыми из известных ископаемых останков тоди являются Palaeotodus emryi, обнаруженные в Вайоминге, которые относят к олигоцену (35 млн лет назад). Американский палеоорнитолог Сторрс Лавджой Олсон в 1976 году полагал, что ближайшим ныне живущим потомком P. emryi является малый момот (Hylomanes momotula), обитающий в Центральной Америке[15]. Предположительные размеры окаменелостей были ближе к малому момоту с пропорционально более коротким и толстым клювом[16]. Из-за бо́льших размеров находки по сравнению с современными тоди Олсон сделал вывод, что ранее род был разнообразнее и включал более крупные виды с более развитыми крыльями[17]. Он считал, что момоты и тоди ранее обитали в Европе и распространились в Северную Америку в начале третичного периода, а в его середине и конце оказались локализованы в Центральной Америке, сохранившей тропический климат. Момоты затем так и остались на континенте, а тоди стали исключительно антильскими видами[15]. Олсон также объяснил более крупные размеры головы тоди: у всех ракшеобразных во время эволюции размер тела изменялся быстрее размера головы, и тоди, как самые маленькие представители отряда, ожидаемо имеют пропорционально большую голову[16].
Более современные находки ископаемых останков тоди в Европе подтверждают теорию Олсона[15]. В 1985 году во Франции были обнаружены останки Palaeotodus escampsiensis (35 млн лет назад) и P. itardiensis (31 млн лет назад). Размеры последнего схожи с размерами найденного ранее P. emryi, в то время как P. escampsiensis по размерам ближе к современным видам. Малое количество костей в этих останках не даёт возможности провести их качественный анализ. Более подробные останки (32 млн лет назад) были обнаружены на юге Германии в 2005 году и предположительно относятся к другому виду Palaeotodus, названному P. itardiensis[17].
Другими факторами, позволяющими считать тоди более близкими к момотам, чем к зимородкам, являются зазубрины на клюве, которые у птенцов сильно похожи на аналогичную структуру у момотов; щетина над клювом; крюк на нижней челюсти у маленьких птенцов; некоторые другие особенности оперения, скелета и мускулатуры. Эволюционные отношения помогает определить также наблюдение за незрелыми птицами[16].
Для выяснения систематики тоди также имеют значения останки Protornis glarniensis, найденные в Швейцарии в 1844 году[17], которые по размеру были меньше малого момота, но больше тоди[16]. Олсон отнёс их к ископаемым момотам[17]. В 1980 году эта гипотеза была поставлена под сомнение: альтернативное мнение высказал американский орнитолог Джоель Кракрафт, который предложил данные останки отнести к Protornis[17].
Основываясь на географическом, палеонтологическом, поведенческом, морфологическом и генетическом анализе, учёные построили предполагаемый сценарий эволюции семейства. Более 30 млн лет назад в Северном полушарии повсеместно обитал гипотетический примитивный Prototodus, который объединял черты Palaeotodus и Protornis. Охлаждение климата, которое произошло в период от 20 до 10 млн лет назад, послужило причиной полного исчезновения Palaeotodus из Европы и резкого сокращения популяции в Северной Америке[16]. Возможно, тоди не сохранились в тропической части Старого Света из-за борьбы с певчими воробьиными птицами за пропитание или из-за активности хищных млекопитающих, усилившейся в связи с тяжёлыми климатическими условиями во время миоцена, которые привели к вымиранию многих видов птиц в Европе[17]. Генетические исследования показали, что семейство тоди отделилось 6—7 млн лет назад. В таком случае дальний предок тоди, всё ещё обладающий длинными крыльями, переселился в Вест-Индию до плейстоценового оледенения, которое случилось миллион лет назад, когда расстояния между островами Центральной Америки и Карибского бассейна были меньше из-за пониженного уровня моря. Ранее считалось, что тоди добрались до Антильских островов в период от 1 миллиона до 10 тысяч лет назад[16].
Эволюция видов
Морфологические особенности разных видов тоди, в особенности оперение и скелет, а также их вокализация, позволяют предположить разделение тоди на две естественные группы. Первая группа включает кубинский, ширококлювый и узкоклювый виды, для которых характерны ярко-розовые бока и соответствующие особенности ухаживания. Ко второй группе относят ямайский и пуэрто-риканский виды, у которых боковые розовые перья отсутствуют полностью либо имеются в крайне небольшом количестве; кроме того, звуки, издаваемые двумя видами во второй группе, почти идентичны по качеству и продолжительности. Вместе с тем последние генетические исследования не поддерживают такое деление[16].
Эволюционные миграционные маршруты тоди проходили с запада на восток, против преобладающих ветров. Предположительно, первым этапом миграции было перемещение по кратчайшему маршруту от Юкатана до Кубы, с последующим распространением до Гаити и Ямайки. Этим же объясняют самое яркое оперение кубинских тоди и их широкую экологическую терпимость. Кроме того, кубинские тоди напоминают предков длиной крыла. Перелёт Никарагуа — Ямайка считается менее вероятным. На Гаити кубинский тоди, судя по интенсивности розового, синего и зелёного цвета в оперении, вокализации и поведению, эволюционировал в ширококлювый вид. По достижении Ямайки кубинский вид превратился в местный эндемик. На связь кубинского и ямайского видов могут указывать сине-серые перья над ухом у последнего, напоминающие сине-голубые пятна кубинского тоди, в то время как у других видов имеется лишь несколько бледно-серых перьев. Кроме того, эти два вида имеют практически равные ширину клюва и длину тела, а также схожее строение крыла[16].
Из-за сильных различий ширококлювого и узкоклювого видов учёные затрудняются сказать, развивались ли они отдельно, когда остров был поделён на две части, или было несколько миграций. Кроме того, есть предположение, что северная и южная популяции узкоклювого тоди в течение последних двух миллионов лет являются эволюционно независимыми. Эти популяции настолько расходятся генетически, что могут даже представлять отдельные виды[16].
Дальнейшая колонизация могла проходить двумя путями: миграция из Гаити в Пуэрто-Рико (узкоклювый вид) или миграция с Ямайки в Пуэрто-Рико. Перемещение, минуя остров Гаити, покрывает большой водный участок и сравнивается с миграцией Dendroica pharetra и Dendroica angelae. Существует и альтернативный вариант, по которому ямайские тоди во время миграции в Пуэрто-Рико поселились на острове Гаити, но позднее там вымерли[16].
Систематика


Учёные относят тодиевых к отряду ракшеобразных, ближайшими родственниками этого семейства являются момотовые и зимородковые[2], при этом момоты, как и тоди, обитают только в Америке, кроме того, они обладают схожим строением клюва[17]. Маленькая птичка todus, предположительно зарянка (Erithacus) или горихвостка (Phoenicurus), упоминается в работах Тита Макция Плавта и Секста Помпея Феста[18]. Французский орнитолог Фредерик де Ла Френе указывает, что в 1725 году английский натуралист Ганс Слоан дал ямайскому тоди название Rubecula viridis elegantissima, а в 1756 году ирландский естествоиспытатель Патрик Броун использовал видовое имя Todus viridis[19] — по всей видимости, основанием для этого стали красное горловое пятно и малые размеры ямайского тоди. Имя Rubecula viridis использовал британский орнитолог Джордж Эдвардс в 1751 году. В современных источниках говорится, что название Todus viridis, также приписываемое Броуну, появилось в 1725 году[18].
Род тоди (Todus) был выделен из зимородков[16] и впервые определён французским зоологом Матюрен-Жаком Бриссоном в 1760 году. В систематике Карла Линнея в 1766 году тоди были расположены рядом с Todirostrum cinereum из семейства тиранновых мухоловок. Теории родства с тиранновыми придерживались французский орнитолог Шарль Люсьен Бонапарт (1850 год), немецкие специалисты Жан Луи Кабанис и Фердинанд Хейн (1860 год), шведский зоолог Карл Якоб Сундеваль (1872 год)[20].
Семейство Todidae было определено ирландским зоологом Николасом Эйлуордом Вигорсом в 1825 году, но включало также золотогузых рогоклювов (Eurylaimus). Немецкие учёные Юлий-Виктор Карус и Карл Эдуард Адольф Герштеккер в 1868 году включили в семейство Platyrhynchus, Euscarthmus и других. Монотипичное семейство, включающее только род тоди, впервые появилось в работе Бонапарта в 1850 году[3].
В 1840 году в работе британского зоолога Джорджа Роберта Грея тодиевые были расположены между сорокопутовыми личинкоедами (Coracina) и рогоклювыми (Eurylaimidae), а в 1848 году он отнёс их к сизоворонковым (Coraciidae). В 1869 году, в более поздней работе, Грей существенно поменял свою точку зрения и расположил тодиевых около момотовых (Momotidae), хотя и продолжал считать их в родстве с рогоклювыми. Ещё до этого, в 1859 году, кубинский натуралист Хуан Гундлах расположил род тоди между зимородками (Alcedo) и настоящими мухоловками (Muscicapa). В 1872 году Джеймс Мери расположил тоди около момотовых и зимородковых, объединив их в группу Serratirostres. Подобное предложение в том же году высказал и британский зоолог Филип Латли Склейтер[20].
В качестве других предполагаемых родственных семейств назывались козодоевые (Caprimulgidae), трогоновые (Trogonidae), якамаровые (Galbulidae), пуховковые (Bucconidae), бородатковые (Capitonidae), котинговые (Cotingidae), манакиновые (Pipridae), цветоедовые (Dicaeidae). В 1882 году было выдвинуто предложение о выделении тоди в отдельный отряд Todiformes, которое, однако, не получило поддержки. В настоящее время отношение семейства к отряду ракшеобразных поддерживается большинством учёных[16].
В 1980 году Кракрафт предложил ограничить тоди и момотов (Todidae + Momotidae) Новым Светом. Однако большая часть современных материалов молекулярного анализа поддерживает теорию клады Todidae + [Momotidae + Alcedinidae], и только один анализ показывают слабую поддержку теории близкого родства таксонов Todidae + Momotidae[17].
Классификация
Родственные связи всех видов тоди никогда не подвергались сомнению, при этом ещё в 1872 году высказывались предположения, что все птицы являются подвидами ямайского тоди[16].
В 1766 году в систематику Линнея вошёл Todus viridis, описание которого, по всей видимости, основывалось на ямайском виде. В дальнейшем под этим именем описывали пуэрто-риканских, ямайских и кубинских тоди. Только в 1837 году британский орнитолог Джон Гульд описал кубинского тоди Todus multicolor как отдельный вид, после чего в 1838 году последовало описание французским натуралистом Рене Примевэром Лессоном пуэрто-риканского тоди Todus mexicanus, экземпляр которого, как считалось, был получен в Мексике. В 1847 году Грей определил четыре вида тоди, включая ширококлювого тоди Todus subulatus, который при этом не был описан (в работе 1848 года того же автора этот вид не учтён). Последовавшие описания рода, предпринятые французскими орнитологами Ла Френе (1847) и Бонапартом (1850), также включали четыре вида. В 1851 году Ла Френе описал узкоклювого тоди Todus anguistirostris на основе экземпляра, предоставленного французским энтомологом Огюстом Салле. В том же году последний высказал предположение, что ширококлювый и узкоклювый экземпляры принадлежат одному виду, а внешние различия объясняются половым диморфизмом. В 1867 году американский орнитолог Спенсер Фуллертон Бэрд пришёл к выводу, что Todus mexicanus обитает на острове Пуэрто-Рико. В работе Грея 1869 года упоминается пять видов тоди[20].
Американский орнитолог Роберт Риджуэй в бюллетене Смитсоновского института 1914 года описывает шесть видов тоди, два из которых — Todus pulcherrimus и Todus todus — обитают на Ямайке[3].
В настоящее время в семействе тодиевых выделяют один род, который включает пять видов[21][22].
| Виды | |||
|---|---|---|---|
| Русскоязычное и научное название |
Изображение | Описание | Распространение |
| Кубинский тоди (Todus multicolor Gould, 1837) |  |
Общая длина — 10—11 см; масса — 4,3—8,5 г. Имеет голубое с боков горло и красно-оранжевое горловое пятно[23]. | Острова Куба, Хувентуд, малые острова на северном побережье Кубы[23]. |
| Узкоклювый тоди (Todus angustirostris Lafresnaye, 1851) |  |
Общая длина — 11 см; масса — 5,2—9,0 г. Характерен более узкий клюв по сравнению со всеми остальными видами[24]. | Западная часть острова Гаити[24]. |
| Ширококлювый тоди (Todus subulatus G. R. Gray, 1847) |  |
Общая длина — 11—12 см; масса — 7,0—10,2 г. Постоянно повторяющийся крик ширококлювого тоди «терп-терп-терп» позволяет отличить его от узкоклювого[25]. | Восточная часть острова Гаити и остров Гонав[25]. |
| Ямайский тоди (Todus todus Linnaeus, 1758)typus |  |
Общая длина — 9—10,8 см; масса самцов — 5,5—7,2 г, самок — 6,4—6,5 г. Самый тихий вид[26]. | Остров Ямайка[26]. |
| Пуэрто-риканский тоди (Todus mexicanus R. Lesson, 1838) |  |
Общая длина — 11 см; масса самцов — 5—7,8 г, самок — 4,8—6,5 г. Самый маленький и наименее яркий вид[27]. | Остров Пуэрто-Рико[27]. |
Примечания
- Бёме Р. Л., Флинт В. Е. Пятиязычный словарь названий животных. Птицы. Латинский, русский, английский, немецкий, французский / Под общ. ред. акад. В. Е. Соколова. — М.: Русский язык, РУССО, 1994. — С. 179. — 2030 экз. — ISBN 5-200-00643-0.
- Burton M., Burton R. Tody // International Wildlife Encyclopedia: Sweetfish - tree snake. — Marshall Cavendish, 2002. — P. 2700—2701. — 3168 p.
- Ridgway R. Superfamily Todi (англ.) // The birds of North and Middle America: a descriptive catalogue, Part VI. — Marshall Cavendish, 1914. — P. 441—449.
- Тоди / Лоскот В. М. // Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов. — М. : Большая российская энциклопедия, 2004—2017.
- HBW Alive: Family Todidae, Morphological Aspects.
- HBW Alive: Family Todidae, Voice.
- HBW Alive: Family Todidae, Habitat.
- HBW Alive: Family Todidae, General Habits.
- Neotropical birds online, Diet and Foraging.
- HBW Alive: Family Todidae, Food and Feeding.
- HBW Alive: Family Todidae, Breeding.
- HBW Alive: Family Todidae, Status and Conservation.
- HBW Alive: Family Todidae, Relationship with Man.
- Todus angustirostris (англ.). The IUCN Red List of Threatened Species.
- Neotropical birds online, Distribution.
- HBW Alive: Family Todidae, Systematics.
- Mayr G., Knopf Ch. W. A tody (Alcediniforme: Todidae) from the early Oligocene of Germany (англ.) // The Auk: Ornithological Advances. — 2007. — Vol. 124, no. 4. — P. 1294—1304.
- Jobling J. A. The Helm Dictionary of Scientific Bird Names. — London: A&C Black Publishers Ltd, 2010. — P. 387. — 432 p. — ISBN 978-1-4081-2501-4.
- de Lafresnaye M. F. Mélanges ornithologiques sur le Todier vert, Todus viridis des auteurs (фр.) // Revue zoologique. — Société cuvierienne, 1847. — Vol. 10. — P. 326—333.
- Sharpe R. B. On the Genus Todus (англ.) // The Ibis, A Quarterly Journal of Ornithology. — 1874. — Vol. 4. — P. 344—355.
- Gill F., Donsker D. & Rasmussen P. (Eds.): Todies, motmots, bee-eaters (англ.). IOC World Bird List (v11.1) (20 января 2021). doi:10.14344/IOC.ML.11.1. Дата обращения: 2 апреля 2021.
- HBW Alive: Family Todidae, Species table.
- Kepler A. K., Kirwan G. M. Cuban Tody (англ.). Handbook of the Birds of the World Alive (10 июня 2018). Дата обращения: 8 сентября 2018.
- Kepler A. K., Kirwan G. M. Narrow-billed Tody (англ.). Handbook of the Birds of the World Alive (11 мая 2018). Дата обращения: 8 сентября 2018.
- Kepler A. K., Kirwan G. M. Broad-billed Tody (англ.). Handbook of the Birds of the World Alive (11 мая 2018). Дата обращения: 8 сентября 2018.
- Kepler A. K., Kirwan G. M. Jamaican Tody (англ.). Handbook of the Birds of the World Alive (11 мая 2018). Дата обращения: 8 сентября 2018.
- Kepler A. K., Kirwan G. M. Puerto Rican Tody (англ.). Handbook of the Birds of the World Alive (11 мая 2018). Дата обращения: 8 сентября 2018.
Литература
- Family Todidae (Todies) / Kepler A. K. // Mousebirds to Hornbills. — Barcelona : Lynx Edicions, 2001. — (Handbook of the Birds of the World : [16 v.] / In: J. del Hoyo, A. Elliott, J. Sargatal ; 1992—2013, vol. 6). — ISBN 978-84-87334-30-6.
Ссылки
- Kepler A. K. Family Todidae (Todies) (англ.). Handbook of the Birds of the World Alive (10 сентября 2014). Дата обращения: 8 сентября 2018.
- Overton L. C. Broad-billed Tody (англ.). Neotropical birds online (2011). Дата обращения: 13 февраля 2018.
| Таксономия |
|---|