Слух млекопитающих
Слух млекопита́ющих — процесс восприятия млекопитающими звуковых волн, их анализа и формирования субъективных ощущений, на основании которых животное реагирует на изменения, происходящие во внешнем мире. За данный процесс у млекопитающих несёт ответственность слуховая сенсорная система, возникшая ещё на раннем этапе эволюции хордовых. Эта система позволяет также воспринимать линейные и угловые ускорения, реализуя тем самым также и чувство равновесия, так что более точное её название — статоакустическая сенсорная система. Её периферическую часть образуют органы слуха и равновесия (уши), промежуточную (обеспечивающую передачу нервных импульсов) — цепи вставочных нейронов, а центральную — слуховые центры в коре головного мозга[1][2].
Распознавание звуковых стимулов у млекопитающих является результатом совместной работы органов слуха и головного мозга. При этом значительная часть акустической информации обрабатывается уже на уровне рецепторов, что позволяет многократно сократить объём такой информации, поступающей к мозгу. Устранение избыточности количества информации неизбежно: если объём акустической информации, поступающей на рецепторы слуховой системы, измеряется десятками тысяч бит в секунду (у человека — порядка 1⋅105 бит/с), то возможности нервной системы по её обработке ограничены десятками бит в секунду[3].
Острота слуха и доступный частотный диапазон у млекопитающих, как правило, весьма значительны, но существенно варьируют у разных видов, причём они сопряжены со теми звуками, которые эти животные способны издавать[4]. Так, человек в норме может слышать звуки в диапазоне от 30 до 17 600 Гц, а у большинства других приматов диапазон воспринимаемых звуков смещён вверх (например, у сенегальского галаго он простирается от 90 до 65 000 Гц, так что эти полуобезьяны способны слышать ультразвуки)[5]. Способностью воспринимать ультразвуки обладают и многие другие млекопитающие (собаки слышат звуки в диапазоне от 67 до 45 000 Гц[6], кошки — от 55 до 78 000 Гц[7]). Особенно велик верхний предел частотного диапазона у видов, использующих эхолокацию (например, у белухи он достигает 123 000 Гц, у некоторых летучих мышей — 110 000 Гц). С другой стороны, усатые киты, жирафы и слоны могут общаться друг с другом инфразвуками, и у них значительно меньшее значение принимает нижний предел частотного диапазона (так, слоны воспринимают звуки в диапазоне от 16 до 12 000 Гц)[5][6][8].
Слух и звуковая сигнализация у млекопитающих обслуживают важнейшие жизненные явления — поиск пищи, распознавание опасностей, опознавание особей своего и чужих видов, коммуникацию между особями, причём различные отряды отличаются разными особенностями слуха. Млекопитающие, хотя и в меньшей мере, чем птицы, обладают звуковой локацией; они издают звуки голосовыми связками и резонируют их воздухоносными путями. Среди издаваемых ими звуков наиболее характерны рёв, стон, мяуканье, шипенье[9][10].
Органы слуха и равновесия
Органы слуха у подавляющего большинства видов млекопитающих хорошо развиты и при этом содержат вестибулярный аппарат[11]. В состав каждого органа слуха входят внутреннее, среднее и наружное ухо (включающее наружный слуховой проход и ушную раковину). Если внутреннее и среднее ухо имеются и у других четвероногих, то наружное ухо представляет собой новоприобретение млекопитающих, облегчающее улавливание звуковых волн. Впрочем, у многих водных и роющих видов ушная раковина вторично исчезает[12][13].
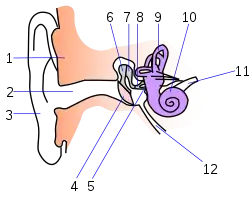
- Наружное ухо
- 1 — череп;
- 2 — слуховой канал;
- 3 — ушная раковина.
- Среднее ухо
- 4 — барабанная перепонка;
- 5 — овальное окно;
- 6 — молоточек;
- 7 — наковальня;
- 8 — стремечко.
- Внутреннее ухо
- 9 — полукружные каналы;
- 10 — улитка;
- 11 — нервы;
- 12 — евстахиева труба.
Наружное ухо
Наружное ухо у млекопитающих устроено достаточно сложным образом. В физическом плане оно представляет собой резонатор (у человека значение собственной резонансной частоты этого резонатора — около 3 кГц)[14]. Ушная раковина армирована хрящом и выполняет роль концентратора звуковых волн, существенно усиливающего тонкость слуха. Обычно ушные раковины подвижны и способствуют достаточной точной (до 6—8°) локализации источников звука в окружающем пространстве. Особенно сильно развита ушная раковина у ночных зверей (например, у летучих мышей), а также у обитающих в лесу копытных, живущих в пустынях псовых и у некоторых других зверей. Наружный слуховой проход защищает от нежелательных внешних воздействий барабанную перепонку, которая представляет собой тонкую мембрану, отделяющую полость среднего уха от слухового прохода и колеблющуюся под воздействием звуковой волны, уловленной ушной раковиной и прошедшей к барабанной перепонке по слуховому проходу[13][15]. Палеонтологи считают, что барабанная перепонка, наличие которой повышает остроту слуха, впервые появилась у непосредственных предков млекопитающих — цинодонтов[16].
Наружный слуховой проход образован хрящом и костной тканью, а его поверхность покрыта тонкой кожей с волосами, в фолликулы которых открываются протоки сальных желёз. Глубже сальных желёз располагаются трубчатые церуминозные железы, выделяющие ушную серу; последняя служит для очистки и смазки слухового прохода, являясь также защитой от бактерий, грибков и насекомых[17].
Среднее ухо
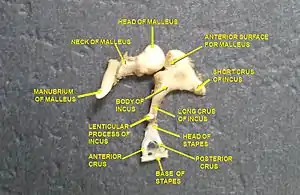
Среднее ухо млекопитающих включает, как и у других четвероногих, барабанную полость, слуховые косточки и евстахиеву трубу; оно выполняет функции аппарата, специализированного для восприятия звуковых волн в воздушной среде[18]. Барабанная полость представляет собой сильно уплощенную и заполненную воздухом камеру, поверхность которой покрыта однослойным эпителием (в основном — плоским, но местами — кубическим или цилиндрическим)[17]. Евстахиева труба, соединяющая полость среднего уха с носоглоткой, отвечает за выравнивание давления по обеим сторонам перепонки[19].
Внутри барабанной полости располагаются три слуховые косточки: стремечко, имеющееся и у других четвероногих, и две специфичные для млекопитающих косточки — молоточек и наковальня (последние гомологичны соответственно сочленовной и квадратной костям прочих четвероногих[20]). Молоточек своим длинным отростком упирается в барабанную перепонку и передаёт колебания последней на наковальню, соединённую, в свою очередь, со стремечком[21][22]. Основание стремечка заходит в закрытое мембраной овальное окно, относящееся уже к внутреннему уху. Три перечисленные косточки составляют в совокупности рычажную систему, которая передаёт колебания перепонки, усиливая их, на мембрану овального окна внутреннего уха. Работа этих косточек заключается в проведении звука из среды с низким сопротивлением (воздух) в среду с высоким сопротивлением (жидкость, которой заполнена улитка) с наименьшими потерями энергии[23]. При этом площадь овального окна значительно меньше площади барабанной перепонки, а потому производимое стремечком на овальное окно давление значительно выше; таким образом, среднее ухо выступает как механический усилитель звуковых волн[18].
К слуховым косточкам прикреплены две мышцы, которые предохраняют структуры внутреннего уха от чрезмерных колебаний при сильном звуке: мышца, напрягающая барабанную перепонку (musculus tensor tympani; связана с молоточком) и стремянная мышца (musculus stapedius; связана со стремечком). При звуке большой интенсивности данные мышцы рефлекторно сжимаются и обеспечивают снижение потока вибраций, проходящего через среднее ухо (акустический рефлекс)[24].
У большинства млекопитающих полость среднего уха прикрывает костная слуховая булла — полая структура, расположенная на внутренней поверхности задней части черепа. Формирование окостеневшей слуховой буллы проходило в нескольких эволюционных линиях млекопитающих независимо — или за счёт выростов окружающих костей, или самостоятельными зонами окостенения[21][25][26].
Внутреннее ухо
Внутреннее ухо находится в каменистой части височной кости и состоит из костного лабиринта и размещённого в нём перепончатого лабиринта. Последний повторяет форму костного лабиринта, но значительно меньше по своим размерам, и заполнен жидкостью — эндолимфой; от костных стенок мембранный лабиринт отделяет другая жидкость — перилимфа. В лабиринте выделяют вестибулярный и слуховой отделы, а также разделяющую их полость неправильной формы — преддверие[27].
В ходе онтогенеза внутреннее ухо закладывается в эктодерме зародыша как плакода, которая превращается сначала в слуховую ямку, а затем — в слуховой пузырёк. Последний делится перетяжкой на две части; верхняя превращается в вестибулярный аппарат, а нижняя — в слуховой отдел внутреннего уха[28].
Слуховой отдел
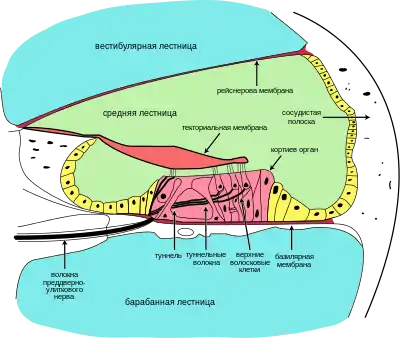
В состав слухового отдела внутреннего уха входит улитка — заполненная жидкостью перепончатая трубка, дугообразно изогнутая (у однопроходных) или спирально завитая (у современных териев — сумчатых и плацентарных; у последних число витков спирали обычно равно двум с половиной, но может доходить и до четырёх с половиной)[29]. Появление спирально завитой улитки отмечено уже у дриолеста (Dryolestes), жившего в позднеюрскую эпоху[30].
По всей длине улитки проходят три параллельных канала: два наружных — вестибулярная лестница) и барабанная лестница — и заключённый между ними более узкий улитковый канал[31][32]. Последний имеет в сечении форму треугольника, стороны которого образованы рейснеровой мембраной, отделяющей улитковый канал от вестибулярной лестницы, сосудистой полоской, образующей наружную стенку канала, и базилярной мембраной — соединительнотканной пластинкой, которая состоит из тонких параллельных коллагеновых волокон различной длины и толщины, натянутых поперёк улиткового канала, и отделяет последний от барабанной лестницы. Внутри улиткового канала располагается также текториальная мембрана — лентовидная пластинка желеобразной консистенции[33].
Все три канала заполнены жидкостями: вестибулярная и барабанная лестницы — перилимфой, а улитковый канал — эндолимфой. Обе жидкости существенно различаются по химическому составу; в частности, в перилимфе высока концентрация ионов натрия Na+, а в эндолимфе — ионов калия К+. Вследствие этого на разделяющих соответствующие каналы мембранах возникает значительная разность электрических потенциалов, что обеспечивает процесс усиления входящих звуковых сигналов энергией[34].
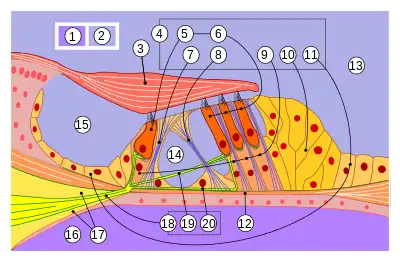
- 1
- перилимфа;
- 2
- эндолимфа;
- 3
- текториальная мембрана;
- 4
- клетки кортиева органа:
- 5,6
- внутренние и внешние волосковые,
- 7,8
- внутренние и внешние столбовые,
- 9
- фаланговые,
- 10
- пограничные,
- 11
- поддерживающие;
- 12
- базилярная мембрана;
- 13
- улитковый канал;
- 14
- кортиев туннель;
- 15
- внутренняя спиральная борозда;
- 16
- барабанная лестница;
- 17
- спиральный лимб;
- 18
- волокна слухового нерва:
- 19
- афферентное,
- 20
- эфферентное
На базилярной мембране располагается кортиев орган — эпителиальное образование, которое тянется вдоль всей улитки, повторяя её ход. Он состоит из двух видов клеток: волосковых клеток, непосредственно отвечающих за восприятие звуковых стимулов, и опорных клеток различного типа, поддерживающих структурную целостность кортиева органа[35][36]. Кортиев орган присущ только млекопитающим; у других четвероногих аналогом кортиева органа служит базилярный сосочек[37][38].
Среди волосковых клеток различают два существенно различных по форме и функциям типа: внутренние и наружные. В типичных случаях внутренние волосковые клетки располагаются на базилярной мембране в один ряд, а наружные — в три и более рядов; эти ряды разделяются опорными клетками[34]. Внутренние волосковые клетки имеют кувшинообразную форму, а наружные — цилиндрическую (у человека общее количество первых составляет 3500, а число вторых может достигать 12 000—20 000)[39].
Вибрации от молоточка, расположенного напротив овального окна, передаются перилимфе. Движущаяся волна перилимфы доходит до конца улитки и заворачивает в барабанную лестницу, откуда движется к основанию улитки и доходит до круглого окна. Энергия этой волны заставляет вибрировать базилярную мембрану, и эта вибрация стимулирует наружные волосковые клетки. Поскольку базиллярная мембрана туго натянута, в каждой её точке наибольший ответ достигается на волны определённой частоты. Каждая волосковая клетка несёт пучок «волосков» (стереоцилий), содержащих внутри актиновые филаменты и погружённых в эндолимфу, заполняющую улитковый канал[40][41].
Вибрирование базилярной мембраны вызывает сдвиг текториальной мембраны и последующее отклонение стереоцилий (таким образом, слуховой рецептор представляет собой механорецептор). Такое отклонение приводит к открытию механочувствительных ионных каналов в мембране волосковой клетки, внутрь клетки входят катионы К+, и клетка деполяризуется. В результате деполяризации открываются кальциевые каналы, внутрь волосковой клетки входит кальций, и она высвобождает нейромедиатор глутамат в синаптическую щель. Это возбуждает афферентный нейрон, и далее сигнал передаётся в ЦНС. Такое возбуждение происходит только при отклонении стереоцилий в одну определённую сторону; это возбуждение сопровождается выделением большего количества глутамата и увеличением частоты генерации потенциала действия афферентным нейроном, однако при отклонении в противоположную сторону имеет место обратный эффект, и клетка гиперполяризуется[40][41].
Вестибулярный аппарат
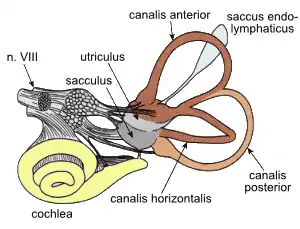
Вестибулярный отдел внутреннего уха, служащий органом равновесия, включает три полукружных канала и два лежащих в преддверии отолитовых органа — круглый мешочек и маточку, иначе называемую эллиптическим мешочком. Полукружные каналы имеют форму изогнутых полукругом трубок, заполнены эндолимфой и локализованы в костных каналах, расположенных в трёх взаимно перпендикулярных плоскостях (наличие именно трёх полукружных каналов присуще вестибулярному аппарату как млекопитающих, так и всех челюстноротых вообще, в то время как у круглоротых число каналов меньше[42]). В месте соединения с маточкой один из концов каждого полукружного канала имеет колбообразное расширение — костную ампулу, а на другом конце такого расширения нет. Маточка, в свою очередь, соединена тонким Y-образным протоком с круглым мешочком[43][44].
В стенке перепончатого лабиринта как в области отолитовых органов, так и в области ампул имеются участки, содержащие сенсорные клетки — механорецепторы[43]. Такие участки применительно к отолитовым органам называются макулами, применительно к ампулам — кристами. В роли механорецепторов в обоих случаях выступают снабжённые пучками волосков волосковые клетки, сходные по устройству с волосковыми клетками слухового отдела. Каждый пучок сверху покрыт купулой — желеобразным веществом, скреплённым фибриллярными волокнами[44][45].
При начале или конце вращательного движения эндолимфа начинает давить на купулу, стремясь сместить её в сторону, противоположную направлению движения, и смещение купулы возбуждает волосковые клетки. Макула каждого из отолитовых органов покрытая отолитовой мембраной, которая содержит множество отолитов, или оокиниев — мелких, но тяжёлых кристаллов карбоната кальция; при этом макула маточки располагается горизонтально, а макула круглого мешочка — вертикально. Под действием силы тяжести или линейного ускорения отолитовые мембраны сдвигаются относительно макул, раздражая расположенные на них волосковые клетки[44][45]. Купулярная система обладает высокой чувствительностью (позволяет фиксировать повороты головы с точностью до 0,005°), но инерционна (характерная постоянная времени лежит в пределах от 10 до 30 с)[46].
Каждая из волосковых клеток вестибулярного аппарата снабжена пучком из нескольких десятков волосков — стереоцилий; на ней также располагается более длинная подвижная ресничка — киноцилия[47]. Если под влиянием механического стимула пучок стереоцилий отклоняется в сторону киноцилии, то волосковая клетка деполяризуется, если от киноцилии — гиперполяризуется. С волосковыми клетками связаны нейроны вестибулярного ядра, активность которых меняется под влиянием нейромедиаторов, выделяемых волосковыми клетками (предположительно глутамата и/или ГАМК). Их аксоны формируют один из корешков преддверно-улиткового нерва, который следует к вестибулярным ядрам. От них информация передаётся в таламус и мозжечок, а из последних — в кору больших полушарий[45].
Промежуточная часть статоакустической сенсорной системы
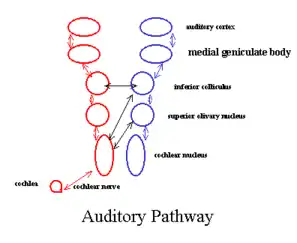
В центральную нервную систему сигнал от улитки поступает по следующему пути: преддверно-улитковый нерв, улитковые ядра продолговатого мозга, верхнее оливное ядро варолиева моста, нижние бугры четверохолмия (лежит в дорсальной части среднего мозга), медиальное коленчатое тело таламуса, слуховая кора (расположена в височной доле коры больших полушарий)[48]. Преддверно-улитковый нерв (VIII пара черепных нервов) состоит из двух ветвей (корешков) — улиткового и вестибулярного нервов: первый обеспечивает передачу акустической информации (в виде электрических импульсов) от волосковых клеток кортиева органа в мозг млекопитающего, а второй отвечает за передачу в мозг информации от аналогичных клеток вестибулярного аппарата[49].
У млекопитающих число нервных волокон, входящих в состав улиткового нерва, исчисляется десятками тысяч (так, у кошки каждое ухо содержит около 50 000 нервных волокон, у человека — около 30 000). Каждую внутреннюю волосковую клетку иннервирует около 20 нервных волокон, причём каждое из последних контактирует с единственной волосковой клеткой. Наружную же волосковую клетку иннервируют от 4 до 6 нервных волокон, каждое из которых одновременно «обслуживает» с десяток волосковых клеток[50]. При этом подходящие к улитке нервные волокна имеют чёткую тонотопическую организацию: у основания улитки располагаются волокна, передающие высокие частоты, в верхушечной области — волокна, передающие низкие частоты[51]. Принцип тонотопической организации нейронных структур, в соответствии с которым нейроны, обрабатывающие звуки с близкими частотными характеристиками, располагаются рядом друг с другом, характерен для всей слуховой системы млекопитающих[52].
Аксоны (центральные отростки) улиткового нерва заканчиваются в продолговатом мозгу — на переднем и заднем улитковых ядрах. На уровне продолговатого мозга формируются разнообразные безусловные рефлексы, возникающие в ответ на звуковые ощущения[53].
В улитковых ядрах берут начало вторые нейроны слухового пути; при этом аксоны нейронов переднего улиткового ядра проходят через трапециевидное тело к верхнему оливному ядру противоположной стороны головы млекопитающего, а аксоны нейронов заднего улиткового ядра идут к верхнему оливному ядру своей стороны[54]. Следовательно, слуховая афферентация от одного рецептора поступает как в противоположное, так и в ипсилатеральное полушарие — особенность слуховой сенсорной системы, отличающая её от других сенсорных систем. Начиная с продолговатого мозга, почти для всех уровней слуховой системы характерен частичный перекрёст слуховых путей, что обеспечивает интегративный характер слуховой афферентации[55].
В верхних оливных ядрах берут своё начало третьи нейроны слухового пути. Их аксоны участвуют в образовании латеральной петли (в состав последней входят также клетки ядра латеральной петли и их отростки), которая состоит из подвергшихся перекрёсту и неперекрещенных слуховых волокон. Заканчиваются эти волокна в подкорковых слуховых центрах — в нижних буграх четверохолмия и медиальном коленчатом теле[56].
Средний мозг выступает как весьма важное звено слуховой системы, поскольку на уровне нижних бугров четверохолмия обрабатывается значительная часть слуховой информации; при этом происходит взаимодействие данных бугров с верхними буграми, обеспечивающее интеграцию слуховой и зрительной афферентаций. В области среднего мозга вновь имеет место частичный перекрёст слуховых путей, за счёт чего часть слуховой информации поступает в противоположное полушарие; именно этот уровень слуховой системы прежде всего обеспечивает бинауральный эффект, позволяя млекопитающему одновременно оценивать и удалённость, и пространственное расположение источника звука (достигается это сопоставлением ощущений, поступающих от левого и правого ушей)[53]. Нижние бугры четверохолмия получают мощную иннервацию со стороны слуховой коры, служа «центральным узлом» между ней и ниже лежащими слуховыми структурами. Они обладают также плотной сетью локальных связей с другими нервными центрами, лежащими в стволе головного мозга; в результате нижние бугры четверохолмия обеспечивают довольно широкий круг функций и, в частности, участвуют в реализации разнообразных сложных рефлексов[57]. Среди таких рефлексов — ориентировочные и сторожевые (настораживание ушей, поворот головы в сторону нового звука)[58].
Что касается медиального коленчатого тела, то оно, будучи частью таламической системы, служит местом дополнительной обработки слуховой информации и её перенаправления на те или иные области слуховой коры[53][59]. Таким образом, именно в нейронах промежуточной части слуховой сенсорной системы выполняется первичная (происходящая на подкорковом уровне) обработка аудиоинформации[60].
Наконец, в подкорковых слуховых центрах берут начало волокна последних нейронов слухового пути, которые ведут в слуховую кору[60].
Суммарное число нейронов, составляющих промежуточную часть акустической сенсорной системы, велико (хотя во много раз меньше, чем число нейронов в центральной части данной системы). У человека оно для каждой стороны головы превышает миллион; из них на долю улитковых ядер приходится примерно 90 000 нейронов, верхнего оливного ядра (включая трапециевидное тело) — 34 000, латеральной петли — 38 000, нижних бугров четверохолмия — 400 000, медиального коленчатого тела — 500 000 нейронов. Наличие столь значительного числа нейронов в подкорковых слуховых центрах говорит о том, что роль промежуточной части акустической сенсорной системы отнюдь не сводится к пассивной передаче сигналов от рецепторных клеток к слуховой коре[61].
Центральная часть статоакустической сенсорной системы
Нейроны коркового центра слуховой сенсорной системы, образующие слуховую кору, располагаются в верхней височной извилине височной доли коры больших полушарий. Этот центр имеет многочисленные ассоциативные связи как с корковыми центрами других сенсорных систем, так и с моторной зоной коры[60]. Среди нейронов слуховой коры есть нейроны, избирательно реагирующие на звуки различной продолжительности, повторяющиеся звуки, шумы с широким частотным диапазоном. В результате происходит объединение информации о высоте, интенсивности тона и временных интервалах между отдельными звуками[48].
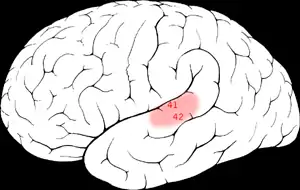
Слуховая кора млекопитающих подразделяется на некоторое число отдельных слуховых полей; часть из них обладает выраженной тонотопической организацией, в других же полях подобная организация выражена слабо. От вида к виду организация слуховой коры может отличаться весьма значительно, так что для обозначения полей в слуховой поре у разных групп млекопитающих используют разные названия (исключением служит слуховая область A1, выделяемая и у однопроходных, и у сумчатых, и у плацентарных[62]). Например, и у приматов, и у хищных выделяют по две первичные слуховые области, одна из которых — A1; вторая у приматов обозначена R, а у хищных — AAF (причём неясно, действительно ли области R и AAF соответствуют друг другу). Аналогично, заднее слуховое поле кошки (P, или PAF) обнаруживает определённое сходство с областями CL и CM у приматов, но остаётся неясным, какие области гомологичны перечисленным у других видов млекопитающих. В слуховой коре летучих мышей, обладающих способностью к эхолокации, найдено несколько областей, связанных с этой способностью и не имеющих очевидных эквивалентов в мозгу приматов[63].
Всего в коре больших полушарий у грызунов выявлено 5—6 слуховых полей, у хищных — 6—9, у мартышковых — 10—12, у человека — свыше 30. Общей чертой организации слуховой коры у млекопитающих является её иерархическое устройство с подразделением на первичную слуховую кору (англ. core region) и окружающие её вторичную слуховую кору (англ. belt region) и третичную слуховую кору (англ. parabelt region). У человека и высших обезьян первичной слуховой коре соответствует 41-е поле Бродмана, вторичной — 42-е и 52-е, третичной — часть 22-го поля[64]. Общее число нейронов в слуховой коре у быка составляет несколько сот тысяч[65], у человека — примерно 100 миллионов[61].
Эхолокация

Некоторые землеройки и морские львы, зубатые киты и в особенности летучие мыши обладают способностью к эхолокации, издавая и воспринимая ультразвуки; у летучих мышей генератором локационных сигналов служат ротовой аппарат и нос, у зубатых китов — система воздушных мешков надчерепного звукового прохода. Землеройки при эхолокации издают импульсы продолжительностью 5—33 мс с частотой 30—60 кГц. Морской лев способен издавать звуки частотой 20—72 кГц, дельфины — 120—200 кГц, что позволяет им определять положение косяков рыб с расстояния до 3 км[4][9]. У летучих мышей ультразвуковая локация почти полностью заменяет слабо развитое зрение[5], однако ни один вид летучих мышей не лишён зрения целиком[66]; более того, зрение используется ими для навигации, особенно на дальних расстояниях, когда радиуса действия эхолокации не хватает[67].
Примечания
- Константинов, Наумов, Шаталова, 2012, с. 35, 335.
- Гистология, цитология и эмбриология, 2004, с. 340—341, 372.
- Воротников, 2005, с. 19—22.
- Дзержинский, Васильев, Малахов, 2014, с. 389.
- Heffner R. S. Primate Hearing from a Mammalian Perspective // The Anatomical Record. Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology. — 2004. — Vol. 281A, № 1. — P. 1111–1122. — doi:10.1002/ar.a.20117. — PMID 15472899.
- Strain G. M. How Well Do Dogs and Other Animals Hear?
- Heffner R. S., Heffner H. E. Hearing Range of the Domestic Cat // Hearing Research. — 1985. — Vol. 19, № 1. — P. 85–88. — doi:10.1016/0378-5955(85)90100-5. — PMID 4066516.
- Vaughan, Ryan, Czaplewski, 2011, p. 27.
- Константинов, Шаталова, 2004, с. 390.
- Vaughan, Ryan, Czaplewski, 2011, p. 40.
- Константинов, Шаталова, 2004, с. 389, 514.
- Гистология, цитология и эмбриология, 2004, с. 372.
- Константинов, Наумов, Шаталова, 2012, с. 335.
- Воротников, 2005, с. 25.
- Дзержинский, Васильев, Малахов, 2014, с. 390—391.
- Черепанов, Иванов, 2007, с. 260.
- Гистология, цитология и эмбриология, 2004, с. 373.
- Дзержинский, Васильев, Малахов, 2014, с. 205.
- Haberman R. S. . Middle Ear and Mastoid Surgery. — New York: Thieme, 2004. — 273 p. — ISBN 1-58890-173-4. — P. 152—154.
- Черепанов, Иванов, 2007, с. 127, 265.
- Дзержинский, Васильев, Малахов, 2014, с. 390.
- Константинов, Шаталова, 2004, с. 389.
- Зильбернагль, Деспопулос, 2013, с. 376.
- Шиффман Х. Р. . Ощущение и восприятие. 5-е изд. — СПб.: Питер, 2003. — 928 с. — ISBN 5-318-00373-7. — С. 183.
- Черепанов, Иванов, 2007, с. 265.
- Кэрролл Р. . Палеонтология и эволюция позвоночных: В 3-х тт. Т. 3. — М.: Мир, 1993. — 312 с. — ISBN 5-03-001819-0. — С. 13—15, 235.
- Гистология, цитология и эмбриология, 2004, с. 374—375, 380.
- Константинов, Шаталова, 2004, с. 513—514.
- Vaughan, Ryan, Czaplewski, 2011, p. 46, 81, 266.
- Luo Zhe-Xi, Ruf I., Schultz J. A., Martin T. Fossil evidence on evolution of inner ear cochlea in Jurassic mammals // Proc. of the Royal Society. B. — 2011. — Vol. 278, № 1702. — P. 28–34. — doi:10.1098/rspb.2010.1148..
- Campbell, 2011, p. 1091.
- Зильбернагль, Деспопулос, 2013, с. 376—377.
- Гистология, цитология и эмбриология, 2004, с. 375—377.
- Kelly M. C., Chen Ping. Development of form and function in the mammalian cochlea // Current Opinion in Neurobiology. — 2009. — Vol. 19, № 4. — P. 395–401. — doi:10.1016/j.conb.2009.07.010. — PMID 4158839.
- Kelly M. C., Chen Ping. Shaping the mammalian auditory sensory organ by the planar cell polarity pathway // The International Journal of Developmental Biology. — 2007. — Vol. 51, № 6—7. — P. 535–547. — doi:10.1387/ijdb.072344mk. — PMID 4158833.
- Гистология, цитология и эмбриология, 2004, с. 377—380.
- Roy D. Development of Hearing in Vertebrates with Special Reference to Anuran Acoustic Communication // Journal of Biosciences. — 1994. — Vol. 19, № 5. — P. 629–644.
- Walsh S. A., Barrett P. M., Milner A. C., Manley G., Witmer L. M. Inner Ear Anatomy is a Proxy for Deducing Auditory Capability and Behaviour in Reptiles and Birds // Proc. of the Royal Society. B. — 2009. — Vol. 276, № 1660. — P. 1355–1360. — doi:10.1098/rspb.2008.1390. Архивировано 24 сентября 2015 года..
- Гистология, цитология и эмбриология, 2004, с. 378.
- Зильбернагль, Деспопулос, 2013, с. 376—379.
- Campbell, 2011, p. 1091—1092.
- Константинов, Шаталова, 2004, с. 514.
- Гистология, цитология и эмбриология, 2004, с. 380.
- Campbell, 2011, p. 1093.
- Ткаченко, 2009, с. 408—412.
- Воротников, 2005, с. 24.
- Гистология, цитология и эмбриология, 2004, с. 381.
- Ткаченко, 2009, с. 406—408.
- Vaughan, Ryan, Czaplewski, 2011, p. 227, 480.
- Pickles J. O. . An introduction to the Physiology of Hearing. 4th ed. — Bingley: Emerald Group Publishing Limited, 2012. — xxiii + 430 p. — ISBN 978-1-78052-166-4. — P. 73.
- Оториноларингология: национальное руководство / Под ред. В. Т. Пальчуна. — М.: ГЭОТАР-Медиа, 2009. — 960 с. — ISBN 978-5-9704-1358-6. — С. 361.
- Хомская, 2011, с. 184.
- Хомская, 2011, с. 185.
- Гистология, цитология и эмбриология, 2004, с. 320, 383—384.
- Хомская, 2011, с. 183.
- Никифоров А. С., Гусев Е. И. . Общая неврология. — М.: ГЭОТАР-Медиа, 2007. — 720 с. — ISBN 978-5-9704-0515-4. — С. 229.
- Malmierca M. S. The Inferior Colliculus: A Center for Convergence of Ascending and Descending Auditory Information // Neuroembryology and Aging. — 2004. — Vol. 3, № 4. — P. 215–229. — doi:10.1159/000096799.
- Лысов, Ипполитова, Максимов, Шевелёв, 2012, с. 64.
- Лысов, Ипполитова, Максимов, Шевелёв, 2012, с. 71.
- Гистология, цитология и эмбриология, 2004, с. 383.
- Warren R. M. . Auditory Perception. An Analysis and Synthesis. 3rd ed. — Cambridge: Cambridge University Press, 2008. — xiii + 264 p. — ISBN 978-0-521-86870-9. — P. 15.
- Karlen S. J., Krubitzer L. The Functional and Anatomical Organization of Marsupial Neocortex: Evidence for Parallel Evolution across Mammals // Progress in Neurobiology. — 2007. — Vol. 82, № 3. — P. 122–141. — doi:10.1016/j.pneurobio.2007.03.003. — PMID 17507143.
- Schnupp J., Nelken I., King A. . Auditory Neuroscience: Making Sense of Sound. — Cambridge, MA: The MIT Press, 2011. — x + 356 p. — ISBN 978-0-262-11318-2. — P. 90—91.
- Hackett T. A. Anatomical Organization of the Auditory Cortex // Journal of the American Academy of Audiology. — 2008. — Vol. 19, № 10. — P. 774–779. — doi:10.3766/jaaa.19.10.5. — PMID 19358457.
- Лысов, Ипполитова, Максимов, Шевелёв, 2012, с. 78.
- Nida Sophasarun. Experts Debunk Bats' Bad Rap. Online extra. National Geographic. Дата обращения: 30 апреля 2013.
- Rachel Kaufman. Bats Crash More When They Rely on Vision (December 15, 2010). Дата обращения 30 апреля 2013.
Литература
- Воротников С. А. . Информационные устройства робототехнических систем. — М.: Изд-во МГТУ им. Н. Э. Баумана, 2005. — 384 с. — ISBN 5-7038-2207-6.
- Гистология, цитология и эмбриология. 6-е изд / Под ред. Ю. И. Афанасьева, С. Л. Кузнецова, H. А. Юриной. — М.: Медицина, 2004. — 768 с. — ISBN 5-225-04858-7.
- Дзержинский Ф. Я., Васильев Б. Д., Малахов В. В. . Зоология позвоночных. 2-е изд. — М.: Издат. центр «Академия», 2014. — 464 с. — ISBN 978-5-4468-0459-7.
- Зильбернагль С., Деспопулос А. . Наглядная физиология. — М.: БИНОМ. Лаборатория знаний, 2013. — 408 с. — ISBN 978-5-94774-385-2.
- Константинов В. М., Наумов С. П., Шаталова С. П. Зоология позвоночных. 7-е изд. — М.: Издат. центр «Академия», 2012. — 448 с. — ISBN 978-5-7695-9293-5.
- Константинов В. М., Шаталова С. П. . Зоология позвоночных. — М.: Гуманитарный издательский центр ВЛАДОС, 2004. — 527 с. — ISBN 5-691-01293-2.
- Лысов В. Ф., Ипполитова Т. В., Максимов В. И., Шевелёв Н. С. . Физиология и этология животных. 2-е изд. — М.: КолосС, 2012. — 605 с. — ISBN 978-5-9532-0826-0.
- Ткаченко Б. И., Брин В. Б., Захаров Ю. М., Недоспасов В. О., Пятин В. Ф. . Физиология человека. Compendium / Под ред. Б. И. Ткаченко. — М.: ГЭОТАР-Медиа, 2009. — 496 с. — ISBN 978-5-9704-0964-0.
- Хомская Е. Д. . Нейропсихология. 4-е изд. — СПб.: Питер, 2011. — 496 с. — ISBN 978-5-459-00730-5.
- Черепанов Г. О., Иванов А. О. Палеозоология позвоночных. — М.: Издат. центр «Академия», 2007. — 352 с. — ISBN 978-5-7695-3104-0.
- Campbell N. A., Reece J. B., Urry L. A. e. a. . Biology. 9th ed. — Benjamin Cummings, 2011. — 1263 p. — ISBN 978-0-321-55823-7.
- Vaughan T. A., Ryan J. M., Czaplewski N. J. . Mammalogy. 5th ed. — Sudbury, Massachusetts: Jones & Bartlett Learning, 2011. — 750 p. — ISBN 978-0-7636-6299-5.
Ссылки
- Чугунов, Антон. Врождённый слуховой аппарат на флексоэлектричестве. // Сайт Biomolecula.ru (27.05.2009). Дата обращения: 18 марта 2018.
- Гутырчик, Алина. Что такое нейропротезирование? Это вредно?. // Сайт Biomolecula.ru (15.08.2017). Дата обращения: 18 марта 2018.