Ренин-ангиотензиновая система
Ренин-ангиотензиновая система (РАС, англ. RAS) или ренин-ангиотензин-альдостероновая система (РААС) — это гормональная система человека и млекопитающих, которая регулирует кровяное давление и объём крови в организме.
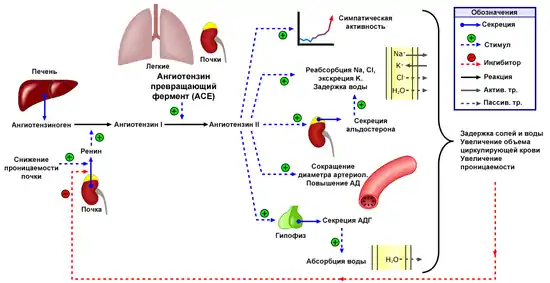
Компоненты системы
Компоненты ренин-ангиотензиновой системы
Ренин-ангиотензин альдестероновый каскад начинается с биосинтеза препроренина на матрице рениновой мРНК в юкстагломерулярных клетках и превращается в проренин путём отщепления 23 аминокислот. В эндоплазматическом ретикулуме проренин подвергается гликозилированию и приобретает 3-D структуру, которая характерна для аспартатных протеаз. Готовая форма проренина состоит из последовательности включающей 43 остатка присоединённых к N-концу ренина, содержащего 339-341 остаток. Предполагается, что дополнительная последовательность проренина (prosegment) связана с ренином для предотвращения взаимодействия с ангиотензиногеном. Большая часть проренина свободно выбрасывается в системный кровоток путём экзоцитоза, но некоторая доля превращается в ренин путём действия эндопептидаз в секреторных гранулах юкстагломерулярных клеток. Ренин, образуемый в секреторных гранулах в дальнейшем выделяется в кровоток, но этот процесс жёстко контролируется давлением, ангиотензином 2, NaCl, через внутриклеточные концентрации ионов кальция. Поэтому у здоровых людей объём циркулирующего проренина в десять раз выше концентрации активного ренина в плазме . Однако, все же остаётся не понятным, почему концентрация неактивного предшественника настолько высока.
Контроль секреции ренина
Активная секреция ренина регулируется четырьмя независимыми факторами:
- Почечным барорецепторным механизмом в афферентной артериоле, который улавливает изменение почечного перфузионного давления.
- Изменениями уровня NaCl в дистальном отделе нефрона. Этот поток измеряется как изменение концентрации Cl- клетками плотного пятна дистального извитого канальца нефрона в области, прилегающей к почечному тельцу.
- Стимуляцией симпатическими нервами через бета-1 адренергические рецепторы.
- Механизмом отрицательной обратной связи, реализованным через прямое действие ангиотензина 2 на юкстагломерулярные клетки.
Секрецию ренина активирует снижение перфузионного давления или уровня NaCl и повышение симпатической активности. Ренин также синтезируется и в других тканях, включая мозг, надпочечник, яичники, жировая ткань, сердце и сосудах.
Контроль секреции ренина — определяющий фактор активности РААС.
Механизм действия ренин-ангиотензиновой системы
Ренин регулирует начальный, ограничивающий скорость, этап РААС путём отщепления N-концевого сегмента ангиотензиногена для формирования биологически инертного декапептида ангиотензина 1 или Ang-(1-10). Первичный источник ангиотензиногена — печень. Долговременный подъём уровня ангиотензиногена в крови, который происходит во время беременности, при синдроме Иценко-Кушинга или при лечении глюкокортикоидами, может вызвать гипертензию, хотя и существуют данные о том, что хроническое повышение концентрации ангиотензина в плазме частично компенсируется снижением секреции ренина.
Неактивный декапептид Ang 1 гидролизуется в клетках эндотелия лёгочных капилляров ангиотензинпревращающим ферментом (АПФ), который отщепляет С-концевой дипептид и, таким образом, формируется октапептид Ang 2 [Ang-(1-8)], биологически активный, мощный вазоконстриктор. АПФ представляет собой экзопептидазу и секретируется главным образом лёгочным и почечным эндотелием, нейроэпителиальными клетками. Ферментативная активность АПФ заключается в повышении вазоконстрикции и снижении вазодилатации.
Ангиотензиноген, синтезированный в печени, преобразуется ренином в ангиотензин 1 (AngI), а затем при участии АПФ — в Ang2. Последний является ключевым звеном РАС, он связывается с ангиотензиновым рецептором первого типа (AT1R). Это взаимодействие вызывает сокращение гладкой мускулатуры бронхов, пролиферацию фибробластов в легких, апоптоз альвеолярных эпителиальных клеток, повышает проницаемость сосудов лёгочной ткани, а также и острый респираторный дистресс-синдром[1]. АПФ2 противодействует активности комплекса ACE-Ang2-AT1R, поскольку гидролизует Ang2 в Angl-7, который уже вызывает снижение артериального давления и стимулирует апоптоз[2].
Новые данные о компонентах ренин-ангиотензиновой системы
Хотя Ang2 наиболее биологически активный продукт РААС, существуют данные, что другие метаболиты агиотензинов 1 и 2 могу также могут иметь значительную активность. Ангиотензин 3 и 4 (Ang 3 & Ang 4) формируются путём отщепления аминокислот с N-конца от Ангиотензина 2 вследствие действия аминопептидаз А и N. Ang 3 и 4 наиболее часто вырабатываются в тканях с высоким содержанием этих ферментов, например, в мозге и почках. Ang 3 [Ang-(2-8)], гептапептид образующий в результате отщепления аминокислоты с N-конца, наиболее часто он встречается в центральной нервной системе, где Ang III играет важную роль в поддержании кров давления. Ang IV [Ang-(3-8)] гексапептид является результатом дальнейшего ферментативного расщепления AngIII. Предполагается, что Ang 2 и 4 работают кооперативно. В качестве примера можно привести повышение кровяного давления в мозгу, вызываемое действием этих ангиотензинов на AT1-рецептор. Причём этот гемодинамический эффект Ang 4 требует наличия как Ang2 так и самого AT1- рецептора. Пептиды, получаемые отщеплением аминокислот с С-конца, могут также иметь биологическую активность. Например, Ang-(1-7), гептапептидный фрагмент ангиотензина 2, может образовываться как из Ang2 так и из Ang1 действием ряда эндопептидаз или действием карбоксипептидаз (например, гомологом АПФ, названным АПФ2) конкретно на Ang2. В отличие от АПФ, АПФ2 не может участвовать в реакции превращения Ang1 в Ang2 и его активность не подавляется ингибиторами ACE (ACEIs). Ang-(1-7) реализующий свои функции через определённые рецепторы, впервые был описан как вазодилататор и как натуральный ингибитор ACEI. Ему также приписываются и кардиопротекторные свойства. АПФ2 может также отщеплять одну аминокислоту с С-конца, результатом такого действия является Ang-(1-9), пептид с неизвестными функциями.
Рецепторы ангиотензина II
Описаны как минимум 4 подтипа рецепторов к ангиотензину.
- Первый тип AT1-R участвует в реализации наибольшего числа установленных физиологических и патофизиологических функций ангиотензина 2. Действие на сердечно-сосудистую систему (вазоконстрикция, повышение давления крови, повышение сократимости сердца, сосудистая и сердечная гипертония), действие на почки (реабсорбция Na+, ингибирование выделения ренина), симпатическую нервную систему, надпочечника (стимуляция синтеза альдостерона). AT1-R рецетор также является посредником во влиянии ангиотензина на клеточный рост, пролиферацию, воспалительные реакции, и оксидативный стресс. Этот рецептор связан с G-белком и содержит семь встроенных в мембрану последовательностей. AT1-R широко представлен во многих типах клеток, являющихся мишенью Ang 2.
- Второй тип AT2-R широко представлен в период эмбрионального развития мозга, почек затем же в период постнатального развития количество этого рецептора падает. Имеются данные, что, несмотря на низкий уровень экспрессии во взрослом организме, AT2 рецептор может выступать в качестве посредника в процессе вазодилятации и также оказывать антипролиферативный и антиапоптотичекие эффекты в гладких мышцах сосудов и угнетать рост кардиомиоцитов. В почках, как предполагается, активация AT2 влияет на реабсорбцию в проксимальных извитых канальцах и стимулировать реакции превращения простагландина E2 в простагландин F2α.2,7. Однако, важность некоторых из этих At2 связанных действий остаётся неизученной.
- Функции третьего типа (AT3) рецепторов не до конца изучены.
- Четвёртый тип рецепторов (AT4) участвует в выделении ингибитора активатора плазминогена (под действием ангиотензина 2, а также 3 и 4). Предполагается, что эффекты характерные для Ang 1-7, включая вазодилятацию, натрийурез, снижение пролиферации, и защита сердца, реализуются через уникальные рецепторы, которые не связываются с Ang 2, такими как MAS рецепторы.
Также нужно отметить, что последние данные указывают на существование высокоаффинных поверхностных рецепторов, которые связывают как ренин, так и проренин. Они находятся в тканях мозга, сердца, плаценты и почек (в поэндотелиальной гладкой мускулатуре и мезангие). Эффекты таких рецепторов направлены на локальное увеличение выработки Ang2 и запуска внеклеточных киназ, таких как, MAP -киназ, к которым относится ERK1 и ERK2. Эти данные пролили свет на Ang2-независимые механизмы клеточного роста, активируемые ренином и проренином.
Влияние на прочие секреции
Как отмечалось ранее Ang2, через AT1 рецепторы стимулирует выработку альдостерона клубочковой зоной надпочечника . Альдостерон наиболее важный регулятор K+- Na+ баланса и таким образом играет важную роль в контроле объёма жидкостей. Он увеличивает реабсорцию натрия и воды в дистальных извитых канальцах и собирательных трубочках (а также в толстой кишке и слюнных и потовых железах) и таким образом вызывает экскрецию ионов калия и водорода. Ангиотензин 2 вместе с внеклеточным уровнем ионов калия - наиболее значимые регуляторы альдостерона, но синтез Ang2 также может быть вызван АКТГ, норадреналином, эндотелином, серотонином, а ингибирован АНП и NO. Также важно отметить, что Ang 2 важный фактор трофики клубочковой зоны надпочечников, которая без его наличия может атрофироваться.
РААС и COVID-19
Группа учёных при изучении механизмов течения коронавирусного заболевания обратила внимание на работу РААС, выявив значительное увеличение концентрации брадикинина под воздействием вируса: он прикрепляется к ангиотензин-рецептору на поверхности клетки и увеличивает синтез АПФ2, попадая с помощью этой молекулы в клетку. Именно повышение концентрации брадикинина (брадикининовый шторм) объясняет многие симптомы пациентов с COVID-19 и вызывает критические осложнения, особенно у пациентов с гипертонией, принимающих препараты с брадикинином для регулирования кровяного давления[3]:
- вызывает неадекватное расширение сосудов=слабость, утомляемость, нарушения ритма сердца;
- увеличивает проницаемость сосудов, что приводит к росту миграции иммунных клеток и усилению воспаления, а также к отёкам и удушью[4];
- усиливает синтез гиалуроновой кислоты (в том числе, в легких), которая вместе с тканевой жидкостью образует гидрогель в просвете альвеол, вызывая проблемы с дыханием и обусловливая неэффективность ИВЛ;
- может увеличивать концентрацию тканевого активатора плазминогена, увеличивая риск кровотечений;
- может приводить к повышению проницаемости гемато-энцефалического барьера, вызывая неврологическую симптоматику[5].
В исследовании отмечается, что основные побочные эффекты ИАПФ — сухой кашель и утомляемость — также обусловлены повышением концентрации брадикинина[3].
См. также
Ссылки
Примечания
- I. Hamming, M. E. Cooper, B. L. Haagmans, N. M. Hooper, R. Korstanje. The emerging role of ACE2 in physiology and disease (англ.) // The Journal of Pathology. — 2007. — Vol. 212, iss. 1. — P. 1–11. — ISSN 1096-9896. — doi:10.1002/path.2162.
- Полина Олеговна Шатунова, Анатолий Сергеевич Быков, Оксана Анатольевна Свитич, Виталий Васильевич Зверев. Ангиотензинпревращающий фермент 2. Подходы к патогенетической терапии COVID-19 // Журнал микробиологии, эпидемиологии и иммунобиологии. — 2020-09-02. — Т. 97, вып. 4. — С. 339–345. — ISSN 2686-7613. — doi:10.36233/0372-9311-2020-97-4-6.
- Шахматова, О.О. Брадикининовый шторм: новые аспекты в патогенезе COVID-19. cardioweb.ru. НАЦИОНАЛЬНЫЙ МЕДИЦИНСКИЙ ИССЛЕДОВАТЕЛЬСКИЙ ЦЕНТР КАРДИОЛОГИИ Министерства здравоохранения РФ. Дата обращения: 23 ноября 2020.
- Huamin Henry Li. Angioedema: Practice Essentials, Background, Pathophysiology (англ.) // MedScape. — 2018-09-04.
- Michael R Garvin, Christiane Alvarez, J Izaak Miller, Erica T Prates, Angelica M Walker. A mechanistic model and therapeutic interventions for COVID-19 involving a RAS-mediated bradykinin storm // eLife. — Т. 9. — ISSN 2050-084X. — doi:10.7554/eLife.59177.