Eciton burchellii
Eciton burchellii (лат., возможное русское название — эцитон бурчелли[3]) — вид средних и мелких по размеру кочевых муравьёв рода Eciton из подсемейства Dorylinae (ранее рассматривался в составе ныне не выделяемого подсемейства Ecitoninae). Постоянных гнёзд не имеют и регулярно совершают массовые кочёвки, перенося с собой всех своих личинок. Один из самых обычных и распространённых по всей Южной и Центральной Америке видов кочевых муравьёв. Встречается от Мексики на севере до Парагвая на юге.
| Eciton burchellii | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 Рабочие и солдат Eciton burchellii | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Без ранга: Тип: Подтип: Надкласс: Класс: Подкласс: Инфракласс: Надотряд: Hymenopterida Отряд: Подотряд: Инфраотряд: Надсемейство: Семейство: Подсемейство: Род: Вид: Eciton burchellii |
||||||||||
| Международное научное название | ||||||||||
| Eciton burchellii (Westwood, 1842)[1] | ||||||||||
| Синонимы | ||||||||||
| ||||||||||

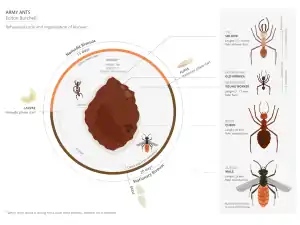

Описание
Мелкие и среднего размера кочевые муравьи. Характерен полиморфизм рабочих. Мелкие рабочие имеют длину 3 мм, наиболее крупные (солдаты) — до 12 мм. Матки и самцы вдвое крупнее — около 2 см. Солдаты желтовато-красные, мелкие рабочие — чёрные с рыжеватым брюшком. Солдаты имеют характерные длинные серповидные жвалы. Усики рабочих и самок 12-члениковые, челюстные щупики 2-члениковые, губные — 3-члениковые. Сложные глаза у рабочих редуцированы, состоят только из одной фасетки. Стебелёк брюшка рабочих состоит из двух сегментов: петиоля и постпетиоля (у маток и самцов из одного только петиоля). Жало развито. Матки бескрылые, простые глазки отсутствуют. Самцы крылатые, их усики 13-члениковые, петиоль отличается вогнутой дорсальной поверхностью.
Каждая колония состоит из одной матки (царицы), расплода (яиц, личинок и куколок) и множества взрослых рабочих. Взрослые рабочие составляют большинство населения. Рабочие делятся на четыре касты: мелкие и средние рабочие, крупные рабочие (субмайоры) и крупные солдаты с огромными челюстями[4][5].
 Солдат сверху
Солдат сверху Голова рабочего
Голова рабочего Рабочий в профиль
Рабочий в профиль
Биология
Данный вид относится к типичным представителям муравьёв-кочевников, которые не строят муравейников, а постоянно перемещаются от одного временного гнезда (бивуака) к другому. При этом они переносят с собой всех личинок. В период остановок, брюшко матки резко увеличивается в размере и она откладывает более сотни тысяч яиц за пару недель. Колонии могут быть большими, включающими от 100 000 до 600 000 взрослых особей. Молодая матка обычно спаривается с 10—20 самцами, в результате колония состоит из соответствующего количества родственных линий рабочих, так что рабочие, принадлежащие к одной и той же линии, являются полными сиблингами, а принадлежащие к разным линиям — нет[6]. Когда размер колонии достигает максимума, это может привести к разделению колонии, когда старая матка возглавляет одну часть семьи, а её дочь возглавляет другую[7].
Фуражировка
Жизнь семьи Eciton burchellii состоит из чередования двух фаз: кочевой и оседлой (когда колония образует «бивуак»). В течение двадцатидневной оседлой фазы развиваются куколки и только что отложенные маткой яйца, и колония совершает фуражировочные рейды примерно через день. После появления новых личинок из яиц и молодых рабочих из куколок, начинается кочевая фаза: в течение пятнадцати дней колония совершает рейды ежедневно[8]. В каждом рейде участвует приблизительно одна треть колонии, то есть до 200 000 муравьёв. В качестве корма используются практически все попадающиеся на пути насекомые, многоножки, паукообразные, а также мелкие позвоночные (ящерицы и другие). Рейды захватывают территорию до 20 метров в ширину и 200 метров в длину[9], причём никогда не занимают одну и ту же область дважды, поэтому тропа к каждому новому местоположению всегда меняется[10]. Колонии E. burchellii избегают мест, где они сами или другие колонии уже фуражировали. Таким образом они гарантируют наличие пропитания в своей области фуражировки и избегают конфликтов с другими представителями своего вида[11].
Влияние вырубки леса
Расчистка тропических лесов наносит ущерб колониям Eciton burchellii. Поскольку они собирают пищу на земле, они чрезвычайно чувствительны к вырубке лесов и разрушению среды обитания[12]. Открытая местность отличается от леса более сильными колебаниями температуры на уровне земли, воздействием прямых солнечных лучей и лучшей видимостью. Для всех вообще насекомых серьёзную угрозу представляет обезвоживание из-за их большого отношения площади поверхности тела к его объёму. Выживание в открытых условиях, вызванных расчисткой леса, для них физиологически затруднено[13]
Было обнаружено, что для муравьёв, живущих в горах, вырубка лесов не так опасна из-за пониженной температуры и большей облачности[14]. Однако количество наземных рейдов уменьшается с увеличением высоты[13].
Влияние феромонов и погоды
В целом, колонии Eciton burchellii не следуют по тропам рейдов предыдущего дня на основе следовых феромонов. Было показано, что дожди задерживают движение муравьёв, иногда на много дней. Проливные дожди изменяют схемы движения фуражиров[15]
Гнёзда
В отличие от других видов общественных перепончатокрылых, муравьи не могут активно регулировать температуру гнезда при помощи таких процессов, как испарение или вентиляция. Поэтому температура в гнезде определяется его местоположением и архитектурой. Для часто мигрирующих видов муравьёв, таких как Eciton burchellii, местоположение гнезда может быть наиболее важным инструментом терморегуляции. Однако E. burchellii не строит физического гнезда. Вместо этого E. burchellii строит живое гнездо из отдельных членов колонии (называемое бивуаком). Терморегуляция внутри этих бивуаков осуществляется путём открытия или закрытия отверстий для воздуха. Члены колонии также могут перестраивать бивуак так, чтобы избежать дождя или прямых солнечных лучей. Обычно бивуак можно найти в полых брёвнах, в норах животных или висящим на деревьях. Во время кочевой фазы муравьи перемещают бивуак на новое место[14]. В среднем колонии требуется 6-8 часов для перемещения своего бивуака. Это происходит в основном ночью[15].
Генетика и биохимия
Масса геномной ДНК равна 0,27 пикограмм[16][17].
В составе мандибулярных желёз в качестве феромона тревоги обнаружено вещество 4-метил-3-гептанон (C8H16O)[18].
Мирмекофилы
Было обнаружено, что в общей сложности 557 различных видов животных каким-либо образом ассоциированы с E. burchellii — больше, чем для любого другого вида животных в мире. На самом деле, зарегистрировано даже большее количество видов, связанных с муравьями, но пока не все они идентифицированы. Хотя некоторые из ассоциаций могут быть случайными, считается, что выживание около 300 видов зависит от E. burchellii в той или иной степени[19]. Существует целая группа так называемых «муравьиных птиц» («ant following birds», дословно — птицы, следующие за муравьями), такие как типичные муравьеловковые, древолазовые, полосатые муравьеловки и дятлы. Насекомые и другие членистоногие, пытающиеся спастись от E. burchellii, попадают в клювы сидящих и летающих рядом птиц, и некоторые их виды в ходе эволюции приспособились добывать большую часть своей пищи, следуя за колоннами муравьёв[20]. Далее, некоторые бабочки затем испольуют помёт птиц, следующих за муравьями, из которого они пьют жидкое содержимое. Мухи из родов Stylogaster (семейство Conopidae) и Calodexia (Tachinidae) летают во множестве вокруг муравьиных колонн, откладывая яйца (или личинок) на бегущих от них тараканов. Исследование спасающихся от муравьёв тараканов показало, что более половины (50-90 %) заражены этими паразитами. Многочисленные виды жуков-стафилинид, которые имитируют муравьёв, можно найти в бивуаках и в колоннах[19].
Также в бивуаках и в колоннах живут многие клещи. На острове Барро-Колорадо в Панаме у 5 % из около 3 тысяч обследованных рабочих муравьёв были клещи, причём наиболее распространены семейства Scutacaridae и Pygmephoridae. Считается, что клещи, как правило, безопасны для муравьёв, будучи симбионтами, а не паразитами. Скорее всего, они используют хозяев для механической транспортировки или пользуются отходами их жизнедеятельности[21]. В частности, на теле рабочих перемещаются клещи Planodiscus и Trichocylliba[22].
На личинках E. burchellii паразитируют наездники диаприиды Doliopria collegii[23].
Птицы и клептопаразитизм
Огромные скопления кочевых муравьёв привлекают так называемых муравьиных птиц, таких как древолазовые или полосатые муравьеловки, которые используют муравьёв как загонщиков насекомых и других мелких членистоногих[20].
Известно множество видов птиц, которые используют фуражировку Eciton burchellii в качестве источника пищи. Когда колонны муравьёв проползают сквозь лесную подстилку, оттуда выбегает и вылетает множество разнообразных членистоногих, которые затем поедаются птицами, ящерицами, насекомыми и даже некоторыми млекопитающими, которые сопровождают муравьиные колонны[24].
Тем не менее, этот источник пищи может быть непредсказуемым, так как зоны набегов колоний Eciton burchellii постоянно меняются. Было показано, что облигатная муравьиная птица глазчатая муравьянка (Phaenostictus mcleannani) использует своеобразную сеть особей для обнаружения колонн муравьёв. Каждая пара этих птиц имеет собственный участок, который она полностью контролирует и который является частью большей, доступной и другим парам, кормовой территории. Благодаря этому, одна семейная пара может отслеживать несколько колоний в день, пользуясь «коллективным разумом». Крики птиц, отлетающих от колонны муравьев, могут быть использованы другими парами как подсказка, где находится колонна[25]. Тем не менее, было показано, что у отбора родичей мало возможностей влиять на групповое питание муравьиных птиц[26].

Двадцать один вид муравьиных птиц практикует «проверку бивуаков» — специализированное поведение, когда птицы оценивают добывающую активность колоний Eciton burchellii и их текущее местоположение. Благодаря этому птицы тратят меньше времени на отслеживание колоний, которые не добывают корм. Вероятно, при проверке бивуаков муравьиные птицы запоминают правильное местоположение колонии, чтобы вернуться к ней в подходящей фазе фуражировочного цикла[27]
Паразитизм муравьиных птиц означает для муравьёв потери, пропорциональные количеству птиц в стае. Это создаёт серьёзное давление отбора на колонии, ведь членистоногие, собранные птицами, составляют почти 50% пищи, потребляемой муравьями. Чтобы противодействовать клептопаразитизму птиц, муравьи прячут крупную добычу или перетаскивают её скрытно, под лесной подстилкой. Кроме того, они создают вдоль фуражировочной тропы продовольственные тайники, защищённые солдатами[24].
Классификация и этимология
| Филогения | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Источник: Winston et al., 2017 [28] |
Вид был впервые описан в 1842 году британским натуралистом Джоном Вествудом (John Obadiah Westwood; 1805—1893) под первоначальным названием Labidus burchellii Westwood, 1842. Видовое название дано в честь британского ботаника и натуралиста Вильяма Бурчелла (1782—1863), собравшего типовой экземпляр в 1826 году в Бразилии[1].
В 1890 году итальянский мирмеколог Карл Эмери перенёс вид в состав рода Eciton[29]. В дальнейшем в некоторых публикациях из видового эпитета выпадала одна буква i. Это связано с тем, что с конца 1800-х годов двойной «i» был признан таксономистами ненужным, и поэтому исходное название стало «Eciton burchelli» с одним i на конце. Однако последние таксономические правила (МКЗН) более строго придерживаются первоначальной формы[2]. Поэтому в настоящее время название Eciton burchelli признано невалидным[2].
По современным молекулярно-генетическим данным (Winston et al., 2017) вид Eciton burchellii сближают с кладой из таксонов Eciton drepanophorum и Eciton hamatum[28].
Подвиды
По данным сайта Б.Болтона (2019) и сайта AntCat[2]:
- Eciton burchellii burchellii (Westwood, 1842)
- Eciton burchellii foreli Mayr, 1886
- Eciton burchellii parvispinum Forel, 1899
- Eciton burchellii urichi Forel, 1899
Распространение
Встречаются во влажных и тёплых тропических лесах в Центральной и Южной Америке. Один из самых обычных и распространённых по всей Неотропике видов кочевых муравьёв от Мексики на севере до Парагвая на юге[30]. Колонии Eciton burchellii не были обнаружены выше 23,8 градусов северной широты[31].
В культуре
Название «Эцитон бурчелли» вошло в культурный обиход в пьесе Сергея Михалкова, по мотивам которой в 1961 году в Театре им. М. Н. Ермоловой бы поставлен спектакль. Журнал «Огонёк» 17 сентября 1961 опубликовал рецензию, в которой назвал одного из героев разновидностью «эцитона бурчелли», безжалостных кочевых муравьёв: «Они прожорливы и безжалостны как истинные тунеядцы», «Живут на марше, набегами и разбоем»[3].
- 1961 — «Обжалованию не подлежит» («Эцитоны бурчелли») (спектакль по мотивам пьесы Сергея Михалкова в Театре им. М. Н. Ермоловой, режиссёр Л. П. Галлис)[3]
- 1978 — Эцитоны Бурчелли (телеспектакль по мотивам одноимённой пьесы Сергея Михалкова, режиссёр Павел Резников)[32][33][34]
Примечания
- Westwood J. O. Monograph of the hymenopterous group, Dorylides // Westwood J. O. Arcana entomologica; or illustrations of new, rare, and interesting insects (Volume 1, No. 5). — London: W. Smith, 1842. — С. 65—80 (pl. 17—20).
- Bolton B. Eciton burchellii (Westwood, 1842) (англ.). An Online Catalog of the Ants of the World. antcat.org. Дата обращения: 7 июля 2019.
- Павлова Н. Марк Иванович не хочет быть узнанным (рецензия на пьесу С.Михалкова «Обжалованию не подлежит» («Эцитоны бурчелли»)) // «Огонёк» : Журнал. — М.: Издательство «Правда», 17 сентября 1961. — № 38 (1787). — С. 21. (недоступная ссылка)
- Resh V. H., Cardé R. T. Encyclopedia of Insects (англ.). — Academic Press, 2009. — ISBN 978-0-08-092090-0.
- Hölldobler B., Wilson E. O. The Ants (англ.). — Cambridge, Massachusetts: Harvard University Press, 1990. — ISBN 978-0-674-04075-5.
- Schneirla T. C. Army ants. A study in social organization (англ.). — San Francisco: W. H. Freeman & Co., 1971.
- Davies N. B., Krebs J. R., West S. A. An introduction to behavioural ecology (англ.). — 4th. — Oxford: Wiley-Blackwell. — ISBN 978-1-4051-1416-5.
- Rettenmeyer C. W., Rettenmeyer M. E., Joseph J., Berghoff S. M. The largest animal association centered on one species: the army ant Eciton burchellii and its more than 300 associates (англ.) // Insectes Sociaux : journal. — 2010. — Vol. 58, no. 3. — P. 281—292. — doi:10.1007/s00040-010-0128-8.
- Gadau J., Fewell J., Wilson E. O. Organization of Insect Societies: From Genome to Sociocomplexity (англ.). — Harvard University Press, 2009. — P. 442. — ISBN 978-0-674-03125-8.
- Powell S., Franks N. How a few help all: living pothole plugs speed prey delivery in the army ant Eciton burchellii (англ.) // Animal Behaviour : journal. — Elsevier, 2007. — Vol. 73, no. 6. — P. 1067—1076. — doi:10.1016/j.anbehav.2006.11.005.
- Willson S. K., Sharp R., Ramler I. P., Sen A. Spatial movement optimization in Amazonian Eciton burchellii army ants (англ.) // Insectes Sociaux : journal. — 2011. — Vol. 58, no. 3. — P. 325—334. — doi:10.1007/s00040-011-0171-0.
- Berghoff S. M., Kronauer D. J. C., Edwards K. J., Franks N. R. Dispersal and population structure of a New World predator, the army ant (англ.) // Journal of Evolutionary Biology : journal. — 2008. — Vol. 21, no. 4. — P. 1125—1132. — doi:10.1111/j.1420-9101.2008.01531.x. — PMID 18422531.
- Kumar A., O'Donnell S. Elevation and forest clearing effects on foraging differ between surface – and subterranean – foraging army ants (Formicidae: Ecitoninae) (англ.) // Journal of Animal Ecology : journal. — 2009. — Vol. 78, no. 1. — P. 91—97. — doi:10.1111/j.1365-2656.2008.01483.x.
- Soare T. W., Tully S. I., Willson S. K., Kronauer D. J. C., O'Donnell S. Choice of nest site protects army ant colonies from environmental extremes in tropical montane forest (англ.) // Insectes Sociaux : journal. — 2010. — Vol. 58, no. 3. — P. 299—308. — doi:10.1007/s00040-010-0134-x.
- Califano D., Chaves-Campos J. Effect of trail pheromones and weather on the moving behaviour of the army ant Eciton burchellii (англ.) // Insectes Sociaux : journal. — 2010. — Vol. 58, no. 3. — P. 309—315. — doi:10.1007/s00040-010-0140-z.
- Detailed Record for Eciton burchelli [База данных о размерах геномов животных] (англ.). genomesize.com. Дата обращения: 8 июля 2019.
- Tsutsui N. D., Suarez J. C., Spagna A. V., Johnston J. S. The evolution of genome size in ants (англ.) // BioMed Central : Журнал. — 2008. — Vol. 8. — P. 64.
- Lalor P. F., and Hughes W. H. O. Alarm behaviour in Eciton army ants (англ.) // Physiological Entomology : Журнал. — 2011. — Vol. 36. — P. 1—7. — doi:10.1111/j.1365-3032.2010.00749.x.
- Rettenmeyer C. W., Rettenmeyer M. E., Joseph J., Berghoff S. M. The largest animal association centered on one species: The army ant Eciton burchellii and its more than 300 associates (англ.) // Insectes Sociaux : journal. — 2010. — Vol. 58, no. 3. — P. 281. — doi:10.1007/s00040-010-0128-8.
- Willis E. O., Oniki Y. Birds and Army Ants (англ.) // Annual Review of Ecology, Evolution, and Systematics. — Annual Reviews, 1978. — Vol. 9. — P. 243—263. — doi:10.1146/annurev.es.09.110178.001331. — .
- Berghoff S. M., Wurst E., Ebermann E., Sendova-Franks A. A. B., Rettenmeyer C. W., Franks N. R. Symbionts of societies that fission: Mites as guests or parasites of army ants (англ.) // Ecological Entomology : journal. — 2009. — Vol. 34, no. 6. — P. 684. — doi:10.1111/j.1365-2311.2009.01125.x.
- Uropodidae Species Listing Архивировано 7 августа 2010 года.. Biology Catalog. Texas A&M University.
- Loiácono M. S., Margaría C. B., Aquino and D. A. Diapriinae Wasps (Hymenoptera: Diaprioidea: Diapriidae) Associated with Ants (Hymenoptera: Formicidae) in Argentina (англ.) // Psyche : Журнал. — 2013. — Vol. 2013, no. Article ID 320590. — P. 1—11. — ISSN 0033-2615.
- Wrege P. H., Wikelski M., Mandel J. T., Rassweiler T., Couzin I. D. Antbirds parasitizing foraging army ants (англ.) // Ecology. — 2005. — Vol. 86, no. 3. — P. 555—559. — doi:10.1890/04-1133.
- Chaves-Campos J. Ant colony tracking in the obligate army ant-following antbird Phaenostictus mcleannani (англ.) // Journal of Ornithology : journal. — 2010. — Vol. 152, no. 2. — P. 497—504. — doi:10.1007/s10336-010-0607-8.
- Chaves-Campos J., DeWoody J. A. The spatial distribution of avian relatives: do obligate army-ant-following birds roost and feed near family members? (англ.) // Molecular Ecology : journal. — 2008. — Vol. 17, no. 12. — P. 2963—2974. — doi:10.1111/j.1365-294X.2008.03811.x. — PMID 18565030.
- Logan C. J., O'Donnell S., Clayton N. S. A case of mental time travel in ant-following birds? (англ.) // Behavioral Ecology : journal. — 2011. — Vol. 22, no. 6. — P. 1149—1153. — doi:10.1093/beheco/arr104.
- Winston M. E., Kronauer D. J. C., Moreau C. S. Early and dynamic colonization of Central America drives speciation in Neotropical army ants (англ.) // Molecular Ecology : Журнал. — 2017. — Vol. 26, no. 3. — P. 859—870. — doi:10.1111/mec.13846.
- Emery C. Studii sulle formiche della fauna neotropica (итал.) // Bull. Soc. Entomol. Ital. : Журнал. — 1890. — Vol. 22. — P. 38—80 (page 39, в составе Eciton).
- Cardé R. T., Resh V. H. A World of Insects: The Harvard University Press Reader (англ.). — Harvard University Press, 2012. — P. 131—. — ISBN 978-0-674-04619-1.
- O'Donnell S., Kaspari M., Kumar A., Lattke J., Powell S. Elevational and geographic variation in army ant swarm raid rates (англ.) // Insectes Sociaux : journal. — 2010. — Vol. 58, no. 3. — P. 293—298. — doi:10.1007/s00040-010-0129-7.
- Эцитоны Бурчелли (1978). telespektakli.ru. Дата обращения: 8 июля 2019.
- Эцитоны Бурчелли. liveinternet.ru. Дата обращения: 8 июля 2019.
- Эцитоны Бурчелли. Телеспектакль по мотивам пьесы Сергея Михалкова (1978). youtube.com. Дата обращения: 8 июля 2019.
Литература
- Borgmeier T. Die Wanderameisen der neotropischen Region (англ.) // Stud. Entomol. : Журнал. — 1955. — Vol. 3. — P. 1—720 (page 178, see also).
- Schneirla T. C. Army Ants: A Study in Social Organization (англ.). — San Francisco : W. H. Freeman & Co, 1971. — P. 349.
- Powell S., Franks N. R. Caste evolution and ecology: a special worker for novel prey (англ.) // Proceedings of the Royal Society B: Biological Sciences (The Royal Society). : Журнал. — 2005. — Vol. 272, no. 1577. — P. 2173—2180.
Ссылки
- Вид Eciton burchellii. lasius.narod.ru. Дата обращения: 7 июля 2019. Архивировано 17 января 2012 года.
- Eciton burchellii Eciton burchellii. antclub.ru. Дата обращения: 7 июля 2019.
- Кочевые муравьи Eciton burchellii затыкают телами дырявые магистрали. membrana.ru. Дата обращения: 7 июля 2019.
- Bolton B. Eciton burchellii (Westwood, 1842) (англ.). An Online Catalog of the Ants of the World. antcat.org. Дата обращения: 7 июля 2019.
- Johnson Norman F. Eciton burchellii (Westwood, 1842) (англ.). Hymenoptera Online (HOL). hol.osu.edu. Дата обращения: 7 июля 2019.
