Наружная морфология насекомых
Насекомые унаследовали от своих предков сегментарное строение, однако метамерность сегментов была практически утрачена и тело разделено на три тагмы: голову, грудь и брюшко. Признаки примитивного метамерного строения сохраняются только в брюшке. Следует отметить, что видимая нами сегментация не соответствует истинной. Сегменты тела насекомых намертво срастаются между собой, в месте их соединения образуется впячивание внутрь — фрагма, используемое для прикрепления мышц. Для обеспечения же подвижности на каждом сегменте у заднего края имеется мягкий участок — сочленение — для защиты немного втянутый внутрь[1].
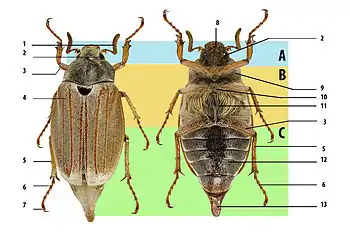
Снаружи тело покрыто выделяемой эпителиальными клетками кутикулой, образующей экзоскелет. Каждый сегмент покрыт хитиновыми щитками, называемыми склериты. В классическом случае выделяют спинные склериты — тергиты, брюшные — стерниты и боковые — плейриты. Согласно теории Снодграсса, плейрит — это уплощившийся и сросшийся со стернитом первый сегмент ноги субкокса, однако зачастую плейриты выделяют и у брюшных сегментов, у которых никаких ног нет. Теория Снодграсса хромает тем, что в природе нет переходных форм, её подтверждающих. Наиболее сильное отступление от классической схемы сегмента наблюдается в голове, где сегменты сильно видоизменены, срослись друг с другом, кроме того снаружи наблюдаются только их тергиты, тогда как стерниты оказываются внутри, образуя так называемый язычок — гипофаринкс[1].
Покровы тела

1: эпикутикула; 1a: цементный слой; 1b: восковой слой; 1с: кутикулиновая пластинка; 1d: белковая эпикутикула.
2: экзокутикула; 3: эндокутикула; 2+3: прокутикула; 4: эпидермис; 5: базальная мембрана; 6: эпидермальная клетка; 6а: восковой канал; 7: железистая клетка; 8: трихогенная клетка; 9: тормогенная клетка; 10: нерв; 11: сенсилла; 12: вырост кутикулы; 13:место, где открывается проток железы.
Кутикула насекомых вырабатывается (секретируется) эпителием, или гиподермой, лежащим на базальной мембране. Эпителий представлен тканью из плотно сомкнутых кубических или цилиндрических эпителиальных клеток и включённых между ними прочих — железистых, трихогенных и других. Эпителиальная ткань, выделяющая кутикулу, также выстилает переднюю и заднюю кишки и трахеи, органы эктодермального происхождения. На кутикуле (особенно эпикутикуле) нередко остаются отпечатки создавших её эпителиальных клеток — так называемый эпителиальный рисунок[1].
Кутикула
Эпителиальные клетки выделяют прочную оболочку — кутикулу. Кутикула формирует наружные покровы насекомых, специальные впячивания в полость тела — фрагмы — для прикрепления мышц, выстилку ппередней и задней кишки и трахей. Кутикула служит для механической защиты, отчасти также для химической защиты, защиты от паразитов ультрафиолетового излучения, для регуляции диффузии, а также, как правило, определяет окраску насекомых. Основное вещество кутикулы — хитин. В нём рассредоточены также инкрусты — структуры другого химического состава[1].
Кутикула состоит из трёх слоёв:
- Внутренний слой кутикулы — эндокутикула — наиболее толстый и прозрачный. Он сложен пластинками, уложенными параллельно поверхности, пронизанными вертикальными каналами. У медведок и жуков слои параллельных балок, слагающих эндокутикулу располагаются под углом друг к другу. Особо прочные покровные органы (надкрылья, пронотум, щитики Pentatomidae и т. д.) укрепляются опорными колоннами.
- Экзокутикула — тонкий, обычно окрашенный, твёрдый за счёт многочисленных белковых и полисахаридных инкруст слой.
- Эпикутикула — очень тонкий слой из воскоподобного вещества кутикулина, вырабатываемого кожными железами. Предохраняет от высыхания наземных членистоногих[1].
Во время линьки под влиянием стероидных гормонов экдизонов, выделяемых проторакальными железами, в гиподерме идёт выработка новой эпикутикулы и активизируются линочные железы. Линочные железы изливают свой секрет между старой эндо- и новой эпи-, от которого эндо- растворяется. Под новой эпи- образуется новая эндо-, вместе с чем растёт проток линочной железы. Раствор старой эндокутикулы либо всасывается гиподермой, либо выпивается, а старые экзо- и эпи- лопаются и остаются в виде шкурки — экзувия. Экзувий тоже можно съесть[1].
Выросты
Складки покровов вместе с гиподермой и полостью тела образуют шипы, рога, гребни, скутеллумы Pentatomidae и т. д. Другая разновидность выростов покровов — хеты. В простейшем случае это комплекс из поры, мембранной клетки, которая образует мембрану в поре и трихогенной клетки, которая образует волосок, торчащий сквозь мембрану. Волосок может как пребывать живым выростом клетки, так и стать омертвевшим кутикулярным образованием. Модификации:
- окружающий пору валик из клеток гиподермы;
- увеличенный вариант (шпора);
- крючья для сцепления крыльев;
- чешуйки бабочек (уплощённый волосок);
- присоски (конец волоскама — диск);
- гидрофобные волоски (для дыхательных трубок водных насекомых);
- ядовитые волоски (в полость волоска входит отросток специальной ядовитой клетки, а сам он хрупок);
- андроконии (аналогично предыдущему, но вместо яда — аттрактант)[1].
Окраска
В зависимости от локализации пигментов окраска тела насекомых может быть:
- кутикулярной — наибольшая посмертная стойкость;
- гиподермальной — зёрна пигментов, локализованные в гиподерме, у некоторых насекомых (палочники) могут перемещаться внутри клетки, меняя цвет покровов по ситуации;
- субгиподермальной — просвечивание внутренностей;
- супракутикулярной — пушки сверху на кутикуле[1].
Кроме непосредственно пигментов окраска может определяться разнообразными оптическими эффектами поверхности (вогнутые чешуйки, призмы, тонкие плёнки). Основной пигмент покровов — меланин — придаёт окраску от жёлтой до чёрной. Обычно чем твёрже участок склерита, тем больше в нём меланинов, тем он, следовательно, темнее. Меланины образуются при окислении хромогенов в присутствии специальных ферментов. Если на теле предполагается рисунок, то он определяется локализацией хромогена, тогда как ферменты распределены по всем покровам. Белые, ярко-жёлтые и оранжевые цвета белянок, ос, сирфид придаются пуриновыми веществами, производными накопленной в куколке экскретной мочевой кислоты. Каротины, флавоны и антоцианы могут заимствоваться насекомыми из поедаемых растений или гемолимфы жертв, заимствовавших пигменты у растений. Зелёный цвет придаётся пигментом биливердином, который окисляясь и восстанавливаясь может менять свой оттенок, что также важно для хамелеонства[2][1].
Голова
Голова (caput) внешне нечлениста, но произошла в результате слияния акрона (головной лопасти) и 5 сегментов в процессе олигомеризации тела. Сохранившимися конечностями этих сегментов являются усики, или антенны первые (antennae) и 3 пары ротовых челюстей — нечленистые верхние челюсти, или мандибулы (mandibulae), членистые нижние челюсти, или максиллы (maxillae) и членистая, внешне непарная нижняя губа (labium), являющаяся второй парой нижних челюстей, слившихся между собой. Вторая пара антенн у насекомых не развивается, но в состав головы входит её сегмент, называемый интеркалярным. Лабрум и три пары челюстей образуют предротовую полость перед ротовым отверстием, где происходит пережёвывание и обработка слюной, а у некоторых сосущих насекомых она является основным насосом. Внутри предротовую полость разделяет на две части стернитный орган — язычок, подглоточник или гипофаринкс[1].
Твёрдой основой головы является черепная коробка (epicranium). На голове различают переднюю поверхность — лоб (frons), который сверху переходит в темя (vertex) и далее назад — затылок (occiput), а с боков в щёки (genae). Обычно лоб очерчен довольно чёткими лобными швами, сливающимися сверху в теменной шов. Спереди ото лба лежит хорошо обособленная пластинка — наличник (clypeus) и далее вперед (вниз) — верхняя губа (labrum), подвижный пластинчатый кожный выступ, прикрывающий сверху ротовые органы. С боков головы расположены сложные глаза (oculi), состоящие из множества зрительных единиц — омматидиев и являющиеся основными органами зрения. Кроме этого, между сложными глазами обычно расположено 1—3 простых глаз, или глазков (ocelli). В зависимости от биологии положение головы неодинаково. Различают гипогнатическую голову (caput hypognathum) — с ротовыми органами, обращёнными вниз, подобно ногам, и прогнатическую голову (caput prognathum) — с ротовыми органами, обращёнными вперёд. Первый тип обычно характерен для растительноядных, а второй — для хищных насекомых. Предельный случай гипогнатической головы — когда ротовые органы смотрят назад — называется опистогнатической головой и характерен для листоблошек и таракановых. Первичной является прогнатическая голова. В процессе её поворота вперёд пятый (лабиальный) сегмент головы оказывается всё дальше ото рта, в связи с чем развивается особый склерит горло (gula), соединяющий 5й сегмент с лабиумом[1].

Усики сидят по бокам лба, между глазами или впереди них, нередко в хорошо обособленной усиковой впадине. Они очень разнообразны, характерны для разных групп насекомых. В своей основе усики состоят из утолщённого основного членика, называемого рукояткой (scapus), за которым следует ножка (pedicellus), и, начиная с третьего членика, располагается основная часть — жгутик (flagellum). Различают несколько типов усиков (см. рис.).
Ротовой аппарат
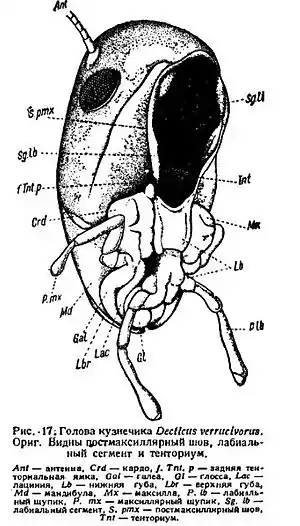
Первичным типом ротового аппарата является ортоптероидный
В ортоптероидном ротовом аппарате лабрум представлен округлой режущей пластинкой, мандибулы мощными нерасчленёнными треугольными пирамидами, максиллы — прямоугольными расчленёнными образованиями из основания (кардо), столбика (стипес) с нижнечелюстным щупиком, наружной лопастинкой (галея) и внутренней лопастинкой (лациния) на конце каждая, лабиум — образованием из сросшихся максиллоподобных челюстей, а потому с двумя нижнегубными щупиками и четырьмя лопастинками.
С переходом к жидкой пище возникают модификации ортоптероидного ротового аппарата. Например, у личинок плавунцов мандибулы длинные острые серповидные и со специальным каналом внутри, предротовая полость зарощена спереди и сообщается с внешним миром только через каналы мандибул. Мандибулами личинка захватывает жертву, впрыскивает в неё пищеварительный сок из средней кишки, а затем теми же каналами всасывает растворённую пищу.
Также с переходом к жидкой пище возникают другие типы ротового аппарата. Все они представляют собой хоботки, различающиеся между собой по строению и по способности к прокалыванию. Неспособные к прокалыванию хоботки называются сосущие и представляют собой разные трубки. Обычно образование трубки связано с удлинением органов, входящих в её состав и редукцией органов, в него не входящих. У пчелы трубка образована длинным стипесами, галеями и лабиумом, причём первые имеют вид уголка в разрезе, а последний плоский. Мандибулы не редуцируются, а используются для строительства и поддержки хоботка.
У чешуекрылых хоботок образован очень длинными галеями, в разрезе имеющими вид полумесяца. Из других органов сохраняются только лабрум и нижнегубные щупики. Внутри головы бабочки имеют специальную мышцу нагоняющую гемолимфу в хоботок для его развёртывания.
У ручейников обнаружены разнообразные хоботки: из галей, из гипофаринкса… Мандибулы обычно остаются. Способные к прокалыванию хоботки называются колюще-сосущими и представляют собой систему из рабочих желобков (для всасывания пищи и введения слюны), прокалывающих стилетов и ножен.
У Nematocera ножны представлены лабиумом, лабрум и гипофаринкс — каналы, а мандибулы и максиллы видоизменены в стилеты.
У це-це (Glossina) и мух жигалок канал образован лабиумом, а прокол осуществляется двумя сверлящими остриями — видоизменёнными нижнегубными щупиками. У блох каналы образованы эпифаринксом и лациниями, ножны — нижнегубными щупиками.
У Hemiptera — ножны представлены лабиумом, лабрум нормальный, щупики отсутствуют, а мандибулы и максиллы превращены в стилеты, слагающие хоботок. Сложенные вместе максиллы образуют два канала, а обнимающие их снаружи максиллы в форме полумесяца в разрезе выполняют прокалывающую функцию.
Особый тип ротового аппарата — мускоидный, характерный для мух. Хоботок образован нижней губой, а её щупики видоизменены в два диска — лабеллума. Каждый лабеллум изрисован 60ю зигзагообразными каналами с круглыми отверстиями, в результате чего получается аппарат с огромной площадью всасывания и способностью к фильтрации[1].
Грудь
Грудь (thorax) состоит из трех сегментов — передне-, средне- и заднегруди (pro-, meso-, metathorax). Тергиты груди называются спинкой (notum), а стерниты — грудкой (sternum). Соответственно 3 сегментам груди различают передне-, средне- и заднеспинку (pro-, meso-, metanotum) и также передне-, средне и заднегрудку (pro-, meso-, metasternum). Каждый плейрит груди подразделяется швом, по крайней мере, на два склерита — спереди эпистерн (episternum) и сзади эпимер (epimerum). Прикрепление к груди органов движения превращает её в локомоторный центр тела, увеличивает размеры за счёт развития мощной мускулатуры и сильному изменению и усложнению описанных выше склеритов.

Ноги
Ноги (pedes) причленены к груди снизу, обычно сидят в тазиковых впадинах и состоят из тазика (coxa), вертлуга (trochanter), бедра (femora), голени (tibia) и лапки (tarsus). Тазик и вертлуг обеспечивают необходимую подвижность ноге. В некоторых случаях вертлуг состоит из двух члеников. Бедро является самой крупной и сильной частью ноги, так как имеет мощную мускулатуру. Его сочленение с голенью называют коленным, а прилегающую к нему часть — коленом (geniculus). Голень по длине примерно равна бедру, но тоньше его, снабжена шипами (spinae), а на вершине — шпорами (calcariae). Лапка обычно расчленена, состоит из 2—5 члеников, на вершине несёт пару коготков (unguiculi), между которыми располагается широкая присоска — аролий (arolium) или узкий эмподий (empodium). Соответственно образу жизни ноги подверглись разнообразной специализации, поэтому различают несколько их типов.
Крылья
Крылья насекомых (alae) представляют собой пластинчатые выросты покровов, связанные с комплексом обслуживающих их мышц и преобразованных склеритов груди. В типичном случае их две пары: передняя, прикреплённая к среднегруди, и задняя — на заднегруди. Крылья состоят из тонкой крыловой пластинки (membrana), укреплённой на твёрдых жилках (venae), играющих роль прочной основы крыла. Жилки делятся на продольные и поперечные, образующие у более низкоорганизованных групп густую сеть и многочисленные ячейки (cellulae). Поперечные жилки у высших форм сокращаются в числе и иногда исчезают совсем, так как главную поддерживающую роль несут продольные, попеременно выпуклые и вогнутые жилки. Поэтому продольные жилки, хотя и претерпели у высших групп значительную эволюцию, всё же сохранили свою основу.
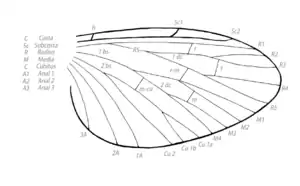
В исходном состоянии различают следующие продольные жилки: костальная (costa или C); субкостальная (subcosta, Sc); радиальная (radius, R), снабжённая сзади ветвью — радиальным сектором (radii sector, RS); срединная, или медиана (media, M), иногда делится на переднюю (MA) и заднюю (MP) ветви; кубитальная (cubitus, Cu), в числе двух, нередко сближенных жилок — передней (CuA) и задней (CuP); анальные (analis, A) в числе 1—4. В некоторых случаях на задних крыльях позади анальных располагается ещё серия югальных (jugalis, Ju) жилок, входящих вместе с анальными в состав складывающегося веера (vannus) крыла. Между продольными жилками располагается серия полей (campo), называемых по пробегающей спереди жилке: костальное, срединное и т. д. Более крупными подразделениями являются области (regio) крыла: главная — спереди от анальных жилок, анальная — между анальными жилками, югальная — между югальными. Этот исходный план жилкования подвергается существенным модификациям путём образования ряда ветвей или выпадения отдельных жилок, их перемещения, слияния и т. д. Наиболее существенно в эволюции крыльев появление у новокрылых насекомых югальной области, обеспечившей особое складывание крыльев вдоль тела. Однако, в процессе дальнейшей эволюции Neoptera югальная область и способность складывать крылья вдоль тела неоднократно утрачивались в разных группах. Также можно отметить происходивший процесс костализации крыла, то есть смещения всех продольных жилок к переднему краю крыла, укреплявшего механическую основу и повышавшего аэродинамические характеристики. Наивысшей степени костализация достигла у перепончатокрылых и двукрылых. Многие отряды пошли по другому пути: у них летательная функция целиком переместилась на задние крылья, а передние превратились в плотные, ороговевшие надкрылья (elytrae), играющие роль покрышек для нежных задних крыльев. В ряде случаев надкрылья полностью утратили жилкование (жуки, кожистокрылые). С совершенствованием полёта от относительной независимости крылья насекомых «переходили» к сцеплению друг с другом различными способами, функционируя как единый орган с перемещением основной нагрузки на первую пару. Другими словами, происходил процесс диптеризации (от лат. названия отряда двукрылых). С дальнейшей эволюцией задняя пара сокращается в размерах, а затем и утрачивается. Возникает высший этап полёта насекомых — морфологическая двукрылость. Всё сказанное определяет выдающуюся роль строения крыльев в классификации и понимании эволюции насекомых.
Брюшко
Брюшко (abdomen) состоит из многих, в целом однотипных, сегментов, исходно из 10, не считая хвостового компонента — тельсона, но в таком виде оно есть лишь у некоторых первичнобескрылых и зародышей. Нередко число сегментов сокращается до 5—6 и менее. На VIII и IX сегментах находятся половые отверстия и очень часто — наружные половые придатки, поэтому эти сегменты обычно называют генитальными. Прегенитальные сегменты у взрослых, как правило, лишены придатков, а постгенитальные имеют развитый лишь тергит X сегмента, тогда как XI сегмент редуцирован и свойственные ему придатки — церки (cerci) переместились на X сегмент. Строение церков весьма разнообразно, а у высших форм они атрофированы. Остатками постгенитальных сегментов являются лежащие дорсально вокруг анального отверстия склериты — сверху анальная пластинка (epiproctus), по бокам и снизу нижние анальные створки (paraproctes). Иногда (тараканообразные, уховёртки) анальной пластинкой называют именно X тергит. Придатками генитальных сегментов являются у самца грифельки (styli) — на IX стерните, у самки — яйцеклад (oviductus) — парные выросты генитальных сегментов, являющиеся видоизмененными конечностями. IX стернит самца образует гипандрий, или генитальную пластинку (hypandrium), но нередко генитальной пластинкой именуют последний видимый стернит вообще, который у самок некоторых Polyneoptera может быть VIII или даже VII. У высших групп все эти части подвергаются редукции или модификации (так, у жалящих перепончатокрылых яйцеклад превращен в жало, хотя используется и по прямому назначению тоже). При редукции настоящего яйцеклада у некоторых групп (например, у многих жуков) возникает вторичный, телескопический яйцеклад из сильно уменьшенных в диаметре вершинных сегментов брюшка.
На конце брюшка самца располагается копулятивный аппарат, имеющий сложное и невероятно разнообразное строение у разных отрядов. Обычно в его составе имеется непарная часть — пенис (penis), имеющий сильно склеротизированную концевую часть — эдеагус (aedeagus). Строение копулятивного аппарата имеет важнейшее значение в систематике, так как его строение заметно различается даже у видов-двойников; часто его изучение позволяет решить труднейшие вопросы классификации родов, семейств и пр.
Литература
- Бей-Биенко Г. Я. Общая энтомология: Учебник для университетов и сельхозвузов. — 3-е изд., доп. — М.: Высшая школа, 1980. — 416 с.
- Шванвич Б. Н. Курс общей энтомологии: Введение в изучение строения и функций тела насекомых (Учебник для гос. университетов) М.—Л.: «Советск. наука», 1949. 900 с.
Примечания
- Шванвич Б. Н.' Курс общей энтомологии: Введение в изучение строения и функций тела насекомых (Учебник для гос. университетов) М.—Л.: «Советск. наука», 1949. 900 с.
- Окраска тела / Словарь — Справочник Энтомолога