Корневые клубеньки
Корневые клубеньки встречаются на корнях растений (преимущественно у семейства бобовых), которые ассоциированы с симбиотическими азотфиксирующими бактериями.
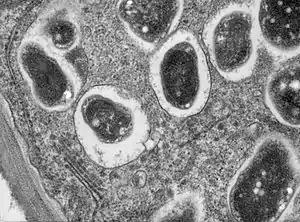
В условиях недостатка азота, растения образуют симбиотическую связь со специфичным для них штаммом клубеньковых бактерий. Такой симбиоз возникал в ходе эволюции независимо несколько раз как внутри семейства бобовых, так и среди других видов, относящихся к кладе розиды[1]. Наиболее изучены клубеньки бобовых, к которым в частности относятся такие важные сельскохозяйственные культуры как фасоль и горох.
В клубеньках бобовых, свободный атмосферный азот восстанавливается до аммония. Который затем ассимилируется, входя в состав органических соединений. При этом образуются аминокислоты (мономеры белков), нуклеотиды (мономеры ДНК и РНК, а также важнейшая молекула обогащённая энергией — АТФ), витамины, флавоны, и фитогормоны.
Способность к симбиотической азотофиксации атмосферного азот делает представителей семейства бобовых идеальной для возделывания сельскохозяйственной культурой, ввиду снижения потребности в азотных удобрения. Более того, высокое содержание доступных для растения форм азота (нитрат NO3− и аммоний NH4+) в почве блокирует развитие клубеньков, поскольку формирование симбиоза для растения становится нецелесообразным.
Энергия для осуществления азотофиксации в клубеньках образуется в результате окисления сахаров (продуктов фотосинтеза), поступающих от листьев. Малат, как продукт распада сахарозы, является источником углерода для симбиотических бактерий.
Процесс фиксации атмосферного азота крайне чувствителен к присутствию кислорода. В связи с чем клубеньки бобовых содержат железосодержащий кислород-связывающий белок — легоглобин. Легоглобин сходен с миоглобином животных, который используется для облегчения диффузии кислорода, использующегося при клеточном дыхании.
Симбиоз
Семейство бобовых
К симбиотической азотофиксации способны многие представители бобовых (Fabaceae): пуэрария, клевер, соевые бобы, люце́рна, люпин, арахис и ройбос. В корневых клубеньках растений имеются симбиотическии ризобии (клубеньковые бактерии). Ризобии продуцируют азотные соединения, необходимые для роста и конкуренции с другими растениями. Когда растение погибает, фиксированный азот высвобождается, делая себя доступным для других растений, тем самым происходит обогащение почвы азотом. Подавляющее большинство бобовых имеют такие образования, однако некоторые (например, Styphnolobium) их не имеют. Во многих традиционных методах ведения сельского хозяйства поля засеваются разными видами растений, и эта смена видов носит цикличный характер. В качестве примера таких растений можно привести клевер и гречиху (не относятся к бобовым, семейство Polygonaceae). Их также принято называть «зелёным навозом».
Ещё одним сельскохозяйственным методом выращивания агрокультурных растений, является их высаживание между рядами деревьев инга. Инга — это небольшое тропическое жестколистное дерево, способное к образованию корневых клубеньков и, соответственно, фиксации азота.
Растения, не относящиеся к семейству бобовых

Несмотря на то, что на сегодняшний день большинство растений, способных к образованию азотфиксирующих корневых клубеньков, относятся к семейству бобовых, существует несколько исключений:
- Parasponia — тропический род семейства коноплёвых, способный к взаимодействию с ризобиями и образованию азотфиксирующих клубеньков;
- Актиноризальные растения, такие как ольха и восковница, также могут образовывать азотфиксирующие клубеньки благодаря симбиотическим отношениям с бактериями Frankia. Эти растения принадлежат к 25 родам, относящимся к 8 семействам.
Способность фиксировать азот распространена в этих семействах не повсеместно. Например, из 122 родов в семействе Розовые, только 4 способны фиксировать азот. Все семейства принадлежат к порядкам Тыквоцветные, Букоцветные и Розоцветные, которые вместе с Бобовоцветными образуют подкласс Розиды. В этом таксоне Бобоцветные были первыми, кто от него ответвились. Таким образом, способность к фиксации азота может быть плезиоморфна и впоследствии могла быть утеряна у большинства потомков исходного азотфиксирующего растения. Однако, возможно, что основные генетические и физиологические предпосылки могли присутствовать и у последнего универсального общего предка всех растений, но реализовались лишь у некоторых современных таксонов.
| Семейство: Род
Берёзовые: Ольха (alders) Коноплёвые: Trema Казуариновые:
|
…… |
Кориариевые: Кориария Датисковые: Датиска |
…… |
…… |
|
…… |
Классификация

На данный момент выделяют два основных типа корневых клубеньков: детерминированные и индетерминированные[2].
Детерминированные корневые клубеньки встречаются у определенных таксонов тропических бобовых, таких как род Glycine (соя), Phaseolus (бобы) и Vigna, а также у некоторых Lotus. Такие корневые клубеньки утрачивают меристематическую активность вскоре после образования, поэтому рост обусловлен лишь увеличением размеров клеток. Это приводит к образованию зрелых клубеньков шаровидной формы. Другие типы детерминированных корневых клубеньков встречаются у многих трав, кустарников и деревьев (например, у арахиса). Они всегда ассоциированы с пазухами боковых или придаточных корней и образуются в результате заражения через повреждения (например, через трещины), в которых образуются эти корни. Корневые волоски при этом в процессе не задействованы. Их внутренняя структура отлична от таковой у соевых бобов[3].
Недетерминированные корневые клубеньки встречаются в большинстве бобовых всех трёх подсемейств как в тропиках, так и в умеренных широтах. Их можно обнаружить у папилиоиноидных бобовых, таких как Pisum (горох), Medicago (люцерна), Trifolium (клевер) и Vicia (вика), а также у всех мимозоидных бобовых, таких как акация, и у цезальпиниоидов. Эти клубеньки получили название «недетерминированных» из-за того, что они их апикальная меристема активна, что приводит к росту клубенька на протяжении всей его жизни. В результате чего формируется клубенёк, имеющий цилиндрическую, иногда разветвлённую форму. Из-за того что они активно растут, можно выделить зоны, которые разграничивают различные стадии развития и симбиоза[4][5][6]:

- Зона I — активная меристема. Здесь формируются новые ткани клубенька, которые затем дифференцируются в другие зоны.
- Зона II — зона инфицирования. Эта зона пронизана инфекционными нитями, состоящими из бактерий. Растительные клетки здесь крупнее, чем в предыдущей зоне, деление клеток останавливается.
- Интерзона II—III — вход бактерий в растительные клетки, содержащие амилопласты. Клетки удлиняются и начинают окончательно дифференцироваться в симбиотические, несущие азотфиксирующие бактерии.
- Зона III — зона фиксации азота. В каждой клетке этой зоны присутствует большая центральная вакуоль и цитоплазма заполнена симбиотическими бактериями фиксирующими азот. Растение наполняет эти клетки легемоглобином, что придаёт им розовый оттенок;
- Зона IV — зона старения. Здесь происходит деградация клеток и их эндосимбионтов. Разрушение гема легемоглобина приводит к появлению зелёного оттенка. Это наиболее изученный тип корневых клубеньков, однако детали различны в клубеньках арахиса и родственных ему растений, а также в клубеньках агрокультурных растений, таких, как люпин. Его клубеньки образуются благодаря прямому заражению ризобиями эпидермы, где инфекционные нити не образуются. Клубеньки растут вокруг корня, образуя структуру наподобие кольца. В этих клубеньках, равно как и клубеньках арахиса, центральная инфицированная ткань однородна. У соевых бобов, гороха и клевера наблюдается недостаток неинфицированных клеток в клубеньках.
Формирование корневого клубенька

Корни бобовых секретируют вещества флавоноиды, которые индуцируют выработку nod-факторов у бактерий. Когда этот фактор распознается корнем, происходит целый ряд морфологических и биохимических изменений: инициируются клеточные деления в корне для создания клубенька, а траектория роста корневого волоска изменяется так, что он обволакивает бактерию вплоть до её полной инкапсуляции. Инкапсулированные бактерии несколько раз делятся, образуя микроколонию. Из этой колонии клетки бактерий входят в развивающийся клубенёк с помощью структуры, называемой инфекционной нитью. Она растёт через корневой волосок вплоть до базальной части клетки эпидермиса, а далее к центру корня. Затем клетки бактерий окружаются мембраной клеток корня растения и дифференцируются в бактериоды, способные фиксировать азот.
Нормальное клубнеобразование занимает приблизительно четыре недели после посадки растения. Размер и форма клубеньков зависит от вида растения, которое было посажено. Так, соя или арахис будут иметь более крупные клубеньки, чем у кормовых бобовых (красный клевер, люцерна). При визуальном анализе количества клубеньков, а также их цвета, учёные могут определить эффективность фиксации азота растением.
Образование клубеньков контролируется как внешними процессами (тепло, рН почвы, засуха, уровень нитратов), так и внутренними (авторегуляция клубнеобразования, этилен). Авторегуляция клубнеобразования контролирует число клубеньков в растении посредством процессов, в которых принимают участие листья. Ткань листа ощущает ранние стадии клубнеобразования через неизвестный химический сигнал, а затем ограничивает дальнейшее развитие клубенька в развивающейся ткани корня. В авторегуляции клубнеобразования участвуют лейцин-богатые повторы (LRR) рецепторных киназ (NARK у соевых бобов (Glycine max); HAR1 у Lotus japonicas, SUNN у Medicago truncatula). Мутации, ведущие к потере функции этих рецепторных киназ ведут к повышенному уровню клубнеобразования. Зачастую аномалии роста корней сопровождаются потерей активности обсуждаемых рецепторных киназ, что указывает на функциональную связь роста клубеньков и корней. Исследование механизмов образований клубеньков показали, что ген ENOD40, кодирующий белок из 12-13 аминокислот, активируется во время клубнеобразования.
Связь со структурой корня
По-видимому, корневые клубеньки у представителей семейства Бобовые образовывались в процессе эволюции минимум три раза и редко встречаются вне этого таксона. Склонность этих растений к развитию корневых клубеньков, скорее всего, связана со структурой корня. В частности, тенденция к развитию боковых корней в ответ на абсцизовую кислоту может способствовать более поздней эволюции корневых клубеньков.
Корневые клубеньки у других видов растений

Корневые клубеньки, которые встречаются у представителей других семейств, таких как параспония — симбиоз с бактериями рода Rhizobium, и те, которые возникают в результате симбиотических взаимодействий с Actinobacteria Frankia, например, у ольхи, значительно отличаются от форм клубеньков, образующихся у бобовых. В симбиозах такого типа бактерии никогда не выходят из инфекционных нитей. Actinobacteria Frankia образует симбиотические отношения со следующими таксонами (семейство указано в скобках): Тыквоцветные (Кориария и Датиска), Букоцветные (Берёзовые, Казуариновые и Восковницевые), Розоцветные (Крушиновые, Лоховые и Розовые). Актиноризальные симбиозы и ризобиальные симбиозы сходны по эффективности фиксации азота. Все эти порядки, включая Fabales, формируют единый азотфиксирующий таксон с более широким таксоном Розиды.
Некоторые грибы формируют клубнеобразные структуры, известные как бугорчатые микоризы, на корнях растений-хозяев. Например, Suillus tomentosus образует такие структуры с сосновой лиственницей (Pinus contorta var. Latifolia). Было показано, что в этих структурах содержатся бактерии, которые способны фиксировать азот. Они фиксируют большой объём азота и позволяют соснам заселять новые территории с бедными почвами[7].
См. также
- Корневые галловые нематоды
Примечания
- Doyle, J. J.; Luckow, M. A. The Rest of the Iceberg. Legume Diversity and Evolution in a Phylogenetic Context (англ.) // Plant Physiology : journal. — American Society of Plant Biologists, 2003. — Vol. 131, no. 3. — P. 900—910. — doi:10.1104/pp.102.018150. — PMID 12644643.
- Martin Crespi; Susana Gálvez. Molecular Mechanisms in Root Nodule Development (неопр.) // Journal of Plant Growth and Regulation. — 2000. — Т. 19, № 2. — С. 155—166. — doi:10.1007/s003440000023. — PMID 11038225. (недоступная ссылка)
- Sprent 2009, Legume nodulation: a global perspective. Wiley-Blackwell
- Fabrice Foucher; Eva Kondorosi. Cell cycle regulation in the course of nodule organogenesis in Medicago (англ.) // Plant Molecular Biology : journal. — 2000. — Vol. 43, no. 5—6. — P. 773—786. — doi:10.1023/A:1006405029600. — PMID 11089876. (недоступная ссылка)
- Hannah Monahan-Giovanelli; Catalina Arango Pinedo; Daniel J. Gage. Architecture of Infection Thread Networks in Developing Root Nodules Induced by the Symbiotic Bacterium Sinorhizobium meliloti on Medicago truncatula (англ.) // Plant Physiology : journal. — American Society of Plant Biologists, 2006. — Vol. 140, no. 2. — P. 661—670. — doi:10.1104/pp.105.072876. — PMID 16384905.
- Willem Van de Velde; Juan Carlos Pérez Guerra; Annick De Keyser; Riet De Rycke; Stéphane Rombauts; Nicolas Maunoury; Peter Mergaert; Eva Kondorosi; Marcelle Holsters; Sofie Goormachtig. Aging in Legume Symbiosis. A Molecular View on Nodule Senescence in Medicago truncatula (англ.) // Plant Physiology : journal. — American Society of Plant Biologists, 2006. — Vol. 141, no. 2. — P. 711—720. — doi:10.1104/pp.106.078691. — PMID 16648219.
- Paul, L.R.; Chapman, B.K.; Chanway, C.P. Nitrogen Fixation Associated with Suillus tomentosus Tuberculate Ectomycorrhizae on Pinus contorta var. latifolia (англ.) // Annals of Botany : journal. — 2007. — Vol. 99, no. 6. — P. 1101—1109. — doi:10.1093/aob/mcm061. — PMID 17468111.
Ссылки
- Корневые клубеньки бобовых в интернет-проекте «Древо жизни»
- Видео и комментарии по корневым клубенькам клевера