Западное внутреннее море
Западное внутреннее море[1] (также Западный внутренний бассейн[2], Западный внутренний пролив[3]; англ. Western Interior Seaway) — древнее эпиконтинентальное море, существовавшее с середины до конца мелового периода на территории современных Канады и США, разделяя её на Ларамидию и Аппалачию.
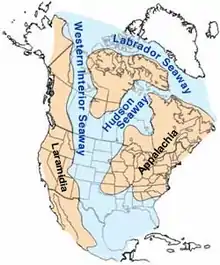
Акватория
Западное внутреннее море было одним из многочисленных шельфовых морей, образовавшихся в результате понижения рельефа дна мирового океана в меловом периоде. Воды с юга (современный Мексиканский залив) и севера соединились в конце альбского яруса. Западное внутреннее море существовало как единая форма рельефа на протяжении почти 35 млн лет, до маастрихтского яруса, завершающего меловой период[4]. Регрессия Западного внутреннего моря началась в конце маастрихта[5].
Вытянутая с севера на юг акватория Западного внутреннего моря граничила с современным Северным Ледовитым океаном на севере и Мексиканским заливом на юге. Западная граница моря проходила вдоль складок рельефа, образованных в ходе Севьерско-Ларамийского орогенеза (современные Скалистые горы)[6]. Море, восточное побережье которого формировала плоская и низменная кратоновая платформа[7] распространялось далеко на восток в сеноманском и туронском ярусе позднего мелового периода, доходя до современных Миннесоты, западного Онтарио и центрального Канзаса[8]. Длина моря, таким образом, составляла порядка 4800 км при максимальной ширине около 1620 км[7]. Бассейн был достаточно мелководным — максимальная глубина в его центральной части, по различным оценкам, достигала от 150 до 300 м[9].
Палеоэкология
На протяжении большей части периода существования Западного внутреннего моря оно располагалось в регионах с умеренным климатом, сменявшимся на субтропический в эпохи наибольшей трансгрессии. Биоты северной части моря (к северу от современных Монтаны, Айдахо и Северной Дакоты) совпадали по большей части с биотами, характерными для побережья тогдашнего Северного океана. В зону мягкого умеренного климата входил регион от северного Вайоминга и Южной Дакоты до Канзаса и севера Юты и Колорадо. Наконец, зона мягкого умеренного климата охватывала территорию к югу до современного Мексиканского залива. На границе между зонами холодного и мягкого умеренного климата жило большинство эндемичных для Западного внутреннего моря видов[7].

В первый, более короткий период соединения южного и северного рукавов Западного внутреннего моря в середине альбского яруса происходило проникновение на север субтропических видов, характерных для тропических рифов. После повторного объединения рукавов движение субтропических видов на север возобновилось, достигнув своего пика в период максимальной трансгрессии в сеноманском ярусе. Резкое снижение солёности морской воды и вызванная взаимной изоляцией слоёв аноксия (снижение уровня растворённого кислорода) привели накануне туронского яруса к массовому вымиранию, известному как сеномано-туронское пограничное биотическое событие, за которым последовали новые вторжения субтропических и тропических организмов в умеренные воды, не достигавшие, однако, прежних масштабов[7]. По отложениям Ниобрарской формации в западном Техасе, сформировавшихся в конце коньякского и сантонском ярусе в южной части Западного внутреннего моря, известны останки птерозавров-никтозавридов, близких к птеранодонам[10].
Западное внутреннее море было богатым на виды беспозвоночных, в частности моллюсков — головоногих, брюхоногих и двустворчатых, о чём свидетельствуют геологические отложения[4]. Широко были представлены также характерные для своей эпохи позвоночные — рыбы, морские рептилии и первые птицы. В отложениях Западного внутреннего моря найдены останки древнейших известных для Северной Америки птиц, относящихся к гесперорнисообразным (Pasquiaornis hardiei и Pasquiaornis tankei из сеноманского яруса), ихтиорнисообразным и, возможно, энанциорнисообразным[11].
В отложениях Западного внутреннего моря найдены останки многочисленных видов рыб — хрящевых (в том числе акул родов Ptychodus, Carcharias и Squalicorax[12]), хрящекостных и костных (последние представлены в том числе гигантскими экземплярами ксифактинов и другими ихтиодектидами, представителями родов Enchodus и Elopopsis, а также древними формами угрей и лососеобразных). Морские рептилии были представлены плезиозаврами (в том числе плиозавридами и эласмозавридами), морскими крокодилами (включая род Teleorhinus, представители которого достигали в длину семи метров) и черепахами[11].
Примечания
- Х. К. Дженкинс. Пелагические фациальные обстановки // Обстановки осадконакопления и фации / Под ред. Х. Рединга. — М.: Мир, 1990. — Т. 2. — С. 131.
- О. Л. Савельева. Орбитально обусловленная ритмичность меловых палеоокеанических отложений // Вестник КРАУНЦ. Науки о Земле. — 2010. — Вып. 16, № 2. — С. 86—95.
- А. Б. Герман. Палеоботаника и климат Земли: взгляд в будущее из геологического прошлого // Вестник Российской академии наук. — 2009. — Т. 79, № 5. — С. 387—396.
- S. He, T. K. Kyser, and W. G. E. Caldwell. Paleoenvironment of the Western Interior Seaway inferred from y18O and y13C values of molluscs from the Cretaceous Bearpaw marine cyclothem // Palaeogeography, Palaeoclimatology, Palaeoecology. — 2005. — Vol. 217. — P. 67—85. Архивировано 4 марта 2016 года.
- Chiarenza A. A., Mannion P. D., Lunt D. J., et al. Ecological niche modelling does not support climatically-driven dinosaur diversity decline before the Cretaceous/Paleogene mass extinction // Nature Communications. — 2019. — Vol. 10. — doi:10.1038/s41467-019-08997-2.
- Andrew D. Miall. Foreland Basin of the Notrh American Western Interior // The Geology of Stratigraphic Sequences. — Springer-Verlag, 1997. — P. 101. — ISBN 3-540-59348-9.
- Erie G. Kauffman. Paleobiogeography and evolutionary response dynamic in the Cretaceous Western Interior Seaway of North America // Jurassic-Cretaceous Biochronology and Paleogeography of North America / G. E. G. Westermann (Ed.). — 1984. — P. 273—306. — (Geological Association of Canada Special Paper 27).
- B. J. Witzke and G. A. Ludvigson. The Dakota Formation in Iowa and the type area // Perspectives on the Eastern Margin of the Cretaceous Western Interior Basin / George W. Shurr, Greg A. Ludvigson, Richard H. Hammond (Eds.). — Boulder, CO, 1994. — P. 43—78. — (Special Paper No. 287 of the Geological Society of America). — ISBN 0-8137-2287-X.
- B. B. Sageman and M. A. Arthur. Early Thuronian paleogeographic/paleobathymetric map, Western Interior, U.S. // Mezozoic Systems of the Rocky Mountain Region, USA / Mario V. Caputo, James A. Peterson and Karen J. Franczyk (Eds.). — RMS-SEPM, 1994. — P. 457—469.
- E. Frey, M.-C. Buchy, W. Stinnesbeck, A. González González, and A. di Stefano. Muzquizopteryx coahuilensis n.g., n. sp., a nyctosaurid pterosaur with soft tissue preservation from the Coniacian (Late Cretaceous) of northeast Mexico (Coahuila). // Oryctos. — 2006. — Vol. 6. — P. 19—39.
- Stephen L. Cumbaa and Tim T. Tokaryk. Recent Discoveries of Cretaceous marine vertebrates on the eastern margins of the Western Interior Seaway // Summary of Investigations, Saskatchewan Geological Survey. — 1999. — Vol. 1. — P. 57—63.
- Liggett, G. A., Shimada, K. E. N. S. H. U., Bennett, C. S., & Schumacher, B. A. Cenomanian (Late Cretaceous) reptiles from northwestern Russell County, Kansas // Paleobios. — 2005. — Vol. 25, № 2. — P. 9—17.
Литература
- Erie G. Kauffman. Paleobiogeography and evolutionary response dynamic in the Cretaceous Western Interior Seaway of North America // Jurassic-Cretaceous Biochronology and Paleogeography of North America / G. E. G. Westermann (Ed.). — 1984. — P. 273—306. — (Geological Association of Canada Special Paper 27).