Teuthowenia megalops
Teuthowenia megalops (лат.) — вид глубоководных кальмаров рода Teuthowenia семейства Кранхииды (Cranchiidae). Встречается в субарктических и умеренных водах северной части Атлантического океана. Это кальмары среднего размера с длиной мантии до 40 см. Особенно крупные глаза стали причиной специфического видового названия (megalops от греческого «большие глаза»). Как и у других представителей рода Teuthowenia отличаются наличием трёх биолюминесцентных органов (фотофоров) на глазных яблоках.
| Teuthowenia megalops | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
.jpg.webp) Teuthowenia megalops | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Тип: Класс: Подкласс: Надотряд: Отряд: Океанические кальмары Семейство: Подсемейство: Taoniinae Род: Вид: Teuthowenia megalops |
||||||||||
| Международное научное название | ||||||||||
| Teuthowenia megalops (Prosch, 1849) | ||||||||||
| Синонимы | ||||||||||
|
||||||||||
| Ареал | ||||||||||
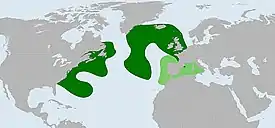
Подтверждённый (тёмно-зелёный) и предполагаемый (светло-зелёный) ареал. |
||||||||||
| Охранный статус | ||||||||||
| ||||||||||

Описание
.jpg.webp)
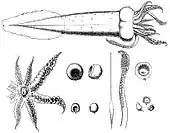
Teuthowenia megalops — кранхииды среднего размера. Характирезуются половым диморфизмом, причём самки более крупные. Мантия достигает длины от 37 до 40 см у самок[4][5][6] и от 18,2 до 24,4 см у самцов[2]. Очень крупные глаза обращены вперёд, каждый из которых обладает тремя биолюминесцентными органами (фотофорами), что является характерной чертой рода Teuthowenia. Один из фотофоров имеет овальную форму, небольшой, расположен в верхней части около зрачка каждого глаза. Два других являются концентрическими и расположены по бокам и снизу глазного яблока. Один из них очень узкий и изогнут на полпути; самый крупный — в форме полумесяца, охватывающего половину нижней поверхности глазного яблока[1][7].
Мантия моллюска коническая и удлинённая с узким задним кончиком. Стенки мантии тонкие и кожистые. Аппарат, запирающий воронку, маленький овальный либо веретенообразный. Присутствует единственный хрящевой бугорок на краях мантии, хотя изредка может вовсе отсутствовать. Длинные и узкие плавники начинаются от 40 до 60% длины мантии, относительно далеко от наиболее широкой части тела. Они проходят вдоль задней части тела, на середине своей длины сливаются друг с другом и заканчиваются парой небольших лепестков[1][2][8]. Щупальца (руки) короткие либо средней длины, примерно от четверти до половины длины мантии. У взрослых самцов и самок третья пара рук самая длинная, за ней следуют вторая, первая и четвёртая пары (фромула длины: III> II> I> IV). Иногда четвёртая и первая пары могут иметь одинаковую или почти равную длину (формула длины: III> II> I = IV)[9]. Взрослые самцы могут иметь первую и вторую пару равной длины (но короче, чем третья пара), или первая пара может быть самой длинной из всех рук (формула длины: III> I = II> IV или I> III> II > IV). Все щупальца имеют два ряда чашеобразных или шаровидных присосок, в которых отсутствуют крючки[8]. Присоски в средней части третьей и второй пары рук значительно увеличены, превышая ширину рук[2].
У молодых особей обоих полов и зрелых самок базальные и срединные присоски рук, как правило, гладкие, хотя маленькие конечные присоски имеют от шести до десяти коротких треугольных зубцов на дистальных и боковых краях. Приблизительно от 6 до 9% кончиков всех рук у молодых особей обоих полов и взрослых самок также превращаются в хватательные конечные органы. У взрослых самцов все щупальца изменены тем или иным образом. Первая пара становится удлинённой и более толстой, с утолщенными средними частями и расширенными защитными мембранами. Вторая пара имеет удлинённые и утолщенные кончики с уменьшенными защитными мембранами. Третья и четвертая пары имеют утонченные кончики. Присоски на концах второй пары имеют кольцеобразные утолщения и обладают мелкими острыми зубцами по всему краю, в то время как у остальных присосок на всех щупальцах маленькие находятся округлые или треугольные зубцы на дистальных и боковых ободках. Средние и базальные присоски третьей и четвёртой пар также имеют кольцеобразные утолщения и значительно уменьшенные отверстия[2].
Ловчие щупальца короткие и мускулистые. На них имеются четыре ряда «запястных» присосок, расположенных зигзагообразно на ½—⅔ от дистального конца щупалец. Концевые присоски расположены на длинных выступах и имеют многочисленные короткие острые крючки на всей кромке. Самые крупные из них имеют от 19 до 24 крючков[2].
Ареал и местообитание
T. megalops — глубоководный пелагический кальмар. Обитает на глубине от 405 до 4515 м при температуре воды от 21 до 3°C, хотя взрослые особи могут изредка встречаться и на глубинах менее 25 м. Кальмары этого вида осуществляют суточную вертикальную миграцию, перемещаясь ближе к поверхности в ночное время и погружаясь глубже днём. Молодь кальмара на планктонной стадии и молодые особи обычно находятся в верхних слоях, около 300 м от поверхности, и постепенно занимают более глубокие слои со взрослением[2][10][11].
Ареал вида ограничен субарктическими и умеренными водами северной части Атлантического океана. В северо-западной Атлантике встречается вдоль восточного побережья Северной Америки, от Лабрадора в Канаде до Джорджии в США; простираясь на восток к Бермудским островам и подводным хребтам Новой Англии[1][12]. В северо-восточной Атлантике обитает между Гренландией и Исландией на севере до Азорских островов на юге за Срединно-Атлантическим хребтом[13][14]; а также в районе Роколлского бассейна, у берегов Великобритании и Ирландии[15][16][17]. Зимой ареал доходит до северной Португалии[18][19]. Один экземпляр также был пойман в Средиземном море, что предполагает, что его ареал, возможно, простирается дальше на юг, чем известно на сегодняшний момент[1]. Вид относительно многочисленный[19][20], а молодые особи часто отлавливаются на мелководье[21].
Ареалы трёх описанных видов Teuthowenia, по-видимому, не перекрываются: Teuthowenia pellucida ограничен южным полушарием, а Teuthowenia maculata встречается в восточной центральной части Атлантического океана у побережья Западной Африки. Таким образом, определение вида может быть сделано на основе места, где он был пойман[22]. Несколько экземпляров кальмара были также обнаружены в Тихом океане в 1969, 1974, 2000 и 2002 годах. Близкое сходство с T. megalops привело к тому, что их постоянно ошибочно идентифицировали как этот вид[23]. Однако по расположению фотофоров на глазах выяснилось, что идентификация была сделана ошибочно[23]. Вероятнее всего, это были Galiteuthis phyllura[24].
Экология и биология
.jpg.webp)
Система флотации
Как и другие члены семейства кранихидов, T. megalops обладают превосходной системой флотации, которая позволяет им оставаться в толще воды при нейтральной плавучести. В их системе флотации используются большие двухлопастные целомические камеры, заполненные хлоридом аммония, образующегося в процессе азотистого обмена. Дело в том, что раствор хлорида аммония имеет более низкую плотность, чем вода, что позволяет кальмару плавать. Экспериментое прокалывание этих камер и выход хлорида аммония приводит к быстрому погружению кальмара. Камера выстлана мышцами, которые регулярно сокращаются и расширяются в перистальтическом режиме. Движение втягивает воду в полость мантии и снова выталкивает её через воронку, что обеспечивает вентиляцию жабер[25][26].
В результате наличия системы флотации T. megalops плохо плавают. В отличие от других кальмаров, им не нужно сокращать свои мантийные мышцы, чтобы дышать, но такая система дыхания ограничивает их способность избегать угроз[26]. Они обычно пассивно плавают в типичной позе «какаду» (поэтому кранихидов также называют «кальмары какаду»). Поза напоминает горизонтального попугая какаду. В этой позе они вращаются вокруг своих веретенообразных пищеварительных желез, единственных внутренних органов кальмара, хорошо видимых через их в основном прозрачные тела. Независимо от направления их головы или мантии пищеварительные железы всегда ориентированы вертикально[21][27].
Жизненный цикл
.jpg.webp)
Жизненный цикл T. megalops недостаточно изучен. Спаривание и нерест происходят на глубинах более 1000 м. На этих глубинах воды тёмные, поэтому биолюминесцентные фотофоры, а также феромоны, выбрасываемые в воду, обеспечивают кальмарам поиск партнёров. Как только самцы находят самок, они засеивают их сперматофорами прямо на мантии. Сперматофоры проникают в мантийную полость самки, где высвобождают сперматозоиды. Самки оплодотворяются единственный раз перед гибелью[22]. Хотя в дикой природе ещё не удалось собрать яйцеклетки у этого вида, анализ собранных беременных самок показывает плодовитость T. megalops в 70—80 тыс. яиц на особь[19].
После вылупления параларва постепенно всплывает к более мелким слоям океана[2][15]. Параларвы отличаются от взрослых тем, что имеют более толстые бочкообразные тела с тупым задним концом. Плавники очень маленькие и не соединённые. Воронка широкая и большая, в отличие от взрослых, на ней нет бугорков на соединении воронки и мантии. Глаза маленькие с видимыми развивающимися фотофорами. Они слегка сплюснуты дорсовентрально и находятся на коротких толстых стебельках. Руки представляют собой простые заглушки длиной от 0,8 до 1,4 мм, когда длина мантии параларвы достигает 8—9 мм. Напротив, щупальца очень длинные и толстые, но без расширенных окончаний. Четыре ряда присосок простираются от окончания почти до основания щупалец. Мантия, голова, руки и щупальца покрыты относительно большими широко расставленными хроматофорами[28].
Параларва формирует характерный бугорок на месте соединения воронки с мантией, когда достигает длины мантии 30—60 мм. Стадия подвзрослости начинается, когда глаза теряют свои стебельки и становятся неподвижными, а размер мантии достигает 75—95 мм. Руки и присоски рук быстро увеличиваются, и у самцов начинают развиваться половые изменения рук (за исключением модификаций первой пары рук, которая происходит почти в зрелости). На этом этапе они, видимо, уже возвращаются на глубину более 1000 м[4].
Продолжительность жизни T. megalops оценивается в 2—3 года[19].
Хищники
На T. megalops охотятся хищные рыбы, такие как синяя акула (Prionace glauca) и меч-рыба (Xiphias gladius)[29]. Эти кальмары также составляют основную часть рациона китообразных, таких как обыкновенная гринда (Globicephala melas), высоколобый бутылконос (Hyperoodon ampullatus), кашалот (Physeter macrocephalus) и клюворыл (Ziphius cavirostris)[19][30].
Защитное поведение
В случае угрозы T. megalops проявляет необычное защитное поведение. Первоначальное проявление защитной реакции этого вида аналогична реакции других головоногих, которые, как правило, выпускают облако чернил в воду и уплывают прочь[31]. Однако, если угроза не исчезает, этот кальмар раздувает себя в шар. Это происходит в несколько этапов и в любое время раздувание может прекратиться, если угроза исчезает[32].
На первом этапе кальмар вывёртывает плавники и гладиус наизнанку. Затем голова и руки убираются в мантийную полость. Щупальца убираютя последними. Наконец, мантия наполняется морской водой, превращая кальмара в идеально сферический объект. В этот момент хроматофоры (обычно простые пятна на в основном прозрачном теле) увеличиваются в четыре раза в диаметре. Они распределяются в концентрические круги на раздутой мантии, сбивая с толку хищника относительно расположения глаз. Если угроза все ещё сохраняется, последним защитным средством является новый выброс чернил. Но, однако, на этот раз чернила высвобождаются не в воду, а в сферическую мантийную полость. Таким образом, кальмар становится полностью непрозрачным чёрным шаром[32].
Кальмар пребывает в защитной форме в течение примерно получаса, после чего начинает постепенно вытягивать сначала голову и щупальца, а затем плавники и гладиус из мантии. Если он уверяется, что угроза исчезла, кальмар постепенно выпускает чернила и морскую воду, хранящиеся в мантии. Всё это происходит без видимой травмы кальмара, несмотря на значительные искажения его тела[32].
Это поведение впервые было обнаружено среди пойманных живых особей в 1972 году морским биологом Питером Ноэлем Дилли. Это стало первым описанием подобного защитного поведения среди головоногих[32]. Позже подобное поведение было обнаружено у других кранхиидов, таких как Cranchia scabra и Teuthowenia pellucida[31][33].
Таксономия и номенклатура
Teuthowenia megalops — один из трёх видов, которые в настоящее время относятся к роду Teuthowenia. Они входят в подсемейство Taoniinae семейства кранхииды (Cranchiidae). Этот вид не имеет широко используемых общих названий. Как и другие роды, принадлежащие к крахниидам, название вида Teuthowenia megalops претерпело многочисленные и запутанные таксономические и номенклатурные пересмотры[1]. Вид в разное время классифицировался под родами Cranchia, Leachia, Megalocranchia и Taonius; а также входил в ныне непринятые роды Desmoteuthis, Hensenioteuthis, Loligopsis и Verrilliteuthis. Различные комбинации родов сочетались со специфическими названиями Cranchia megalops, Leachia hyperborea, Taonidium pfefferi (ныне nomen dubium), Desmoteuthis tenera и Desmoteuthis thori[2].
Таксономическая история
Впервые этот вид был описан датским зоологом Фердинандом Виктором Альфонсом Прошем в 1849 году как Cranchia megalops[34]. Типовые образцы были параларвами, выловленными у Фарерских островов[2]. Прош включил новый вид в новое подсемейство Cranchidæ (ныне семейство Cranchiidae), заметив его сходство с Cranchia scabra и Cranchia maculata (теперь Teuthowenia maculata). Далее он разделил группу на два подрода, поместив последние два в подрод Cranchia и выделив Cranchia megalops в подрод Owenia (названный в честь британского биолога Ричарда Оуэна). Видовое название было получено от греческого «большие глаза»: μέγας (мегас, «великий») и ὤψ (ops, «глаз» или «лицо»)[35].
В 1850 году датский малаколог Отто Андреас Лоусон Мёрч указал на ошибки в первоначальном описании Проша[34]. В 1856 году датский зоолог Япетус Стенструп упомянул образец кальмара-кранхиида, обнаруженный в море Баффина, который он назвал Leachia hyperborea[36]. Позже, в 1861 году, Стенструп основал новый род Taonius, в который включил Loligo pavo (сейчас Taonius pavo) и описанный им вид Leachia hyperborea (как Taonius hyperboreus). В той же работе он также первым указал на то, что Прош допустил ошибку, включив туда молодого Gonatus, определив его как Cranchia megalops[37][38]. В 1879 году американский малаколог Джордж Вашингтон Трион включил Teuthowenia megalops в род Cranchia вместе с Teuthowenia maculata (тогда Cranchia maculata), но выделил Taonius hyperboreus Стенструпа в род Loligopsis вместе с Taonius pavo[39].
В 1881 году американский зоолог Эддисон Эмери Веррил основал род Desmoteuthis для нового вида, названного им Desmoteuthis tenera, пойманного у берегов Новой Англии. Он также обнаружил несколько экземпляров Taonius pavo, которые он неправильно идентифицировал как Taonius hyperboreus, что привело его к включению Taonius hyperboreus в этот новый род как Desmoteuthis hyperborea[40]. В 1884 году, а затем в 1886 году британский зоолог Уильям Эванс Хойл обнаружил ошибку Веррила после изучения хорошо иллюстрированной статьи Веррила. Он синонимизировал род Desmoteuthis назад к Taonius Стенструпа. Он также предварительно определил Desmoteuthis tenera Веррила как возможный синоним Taonius hyperboreus[41][42]. В 1884 году немецкий зоолог Георг Иоганн Пфеффер основал род Megalocranchia, используя типовой вид Megalocranchia maxima. Хотя типовые образцы были утрачены во время Второй мировой войны, исследования его описания и иллюстраций показывают, что Megalocranchia maxima определенно не принадлежали к роду Teuthowenia. Пфеффер синонимизировал Megalocranchia с Desmoteuthis Веррила в 1900 году. Он в основном следовал выводам Хойла относительно Taonius pavo, но сохранил род Desmoteuthis для Taonius hyperboreus. Он также повысил подрод Owenia Проша до уровня рода, переименовав таким образом Cranchia megalops в Owenia megalops[43]. Однако имя Owenia уже было занято родом полихетовых червей, поэтому в 1910 году немецкий зоолог Карл Хун видоизменил род на Teuthowenia, от греч. τευθίς («кальмар») + Owenia, как в предыдущем названии. Хун также впервые описал Desmoteuthis pellucida (сейчас Teuthowenia pellucida) в той же работе[44].
В 1912 году американский зоолог Сэмюэль Стиллман Берри согласился с Хойлом, заключив, что Desmoteuthis Веррила является синонимом Taonius. Но он сделал ошибку, используя Megalocranchia Пфеффера для Desmoteuthis hyperborea; и необъяснимым образом установил новый род Verrilliteuthis для Desmoteuthis tenera (который ранее уже был признан синонимом Desmoteuthis hyperborea)[45]. Пфеффер последовал его примеру в том же году, воскресив Megalocranchia в согласии с Берри, хотя он все ещё сохранил Desmoteuthis hyperborea у Desmoteuthis[46].
В 1925 году немецкий малаколог Эдуард Дегнер впервые описал этапы развития Desmoteuthis hyperborea во время датских океанографических экспедиций (1908—1910) в Средиземное море на борту датского исследовательского судна Thor. Он также описал новый вид Desmoteuthis thori из экземпляра молодой самки кальмара, который имел захватывающие щупальца. Он не смог подключить его к Teuthowenia megalops по двум причинам. Во-первых, ему не хватало образцов Desmoteuthis hyperborea между стадиями роста с длиной мантии от 6 до 84 мм. Во-вторых, он не осознавал, что отличительные конечные органы его экземпляра для Desmoteuthis thori на самом деле были характерными для Teuthowenia megalops, обнаруживаемых только у зрелых и взрослых самок[2]. В 1934 году немецкий зоолог Иоганнес Тиле ещё более усложнил ситуацию, включив Teuthowenia в роду Hensenioteuthis Пфеффера (ныне nomen dubium), вместе с Helicocranchia, Ascoteuthis и номинальным подродом Hensenioteuthis. В то же время он также принял Verbilliteuthis hyperborea[47].
Лишь в 1956 году датский морской биолог Бент Дж. Муус обнаружил, что Cranchia (Owenia) megalops Проша и Leachia hyperborea Стенструпа фактически принадлежат к одному и тому же виду. Первый является стадией параларвы второго. Он синонимизировал их под именем Desmoteuthis megalops, к сожалению, сохранив род Verrill, несмотря на то, что он был синонимичен с Taonius. Как и Хун, он также допустил ошибку, включив в него несколько других не связанных между собой экземпляров, в том числе Helicocranchia pfefferi Анны Л. Масси, Desmoteuthis pellucida и Teuthowenia antarctica Хуна (в настоящее время известные как Galiteuthis glacialis); таким образом, приходя к ошибочному выводу, что Teuthowenia megalops обитают как в северном, так и в южном полушариях[48][49]. В 1960 году американский теутолог Гилберт Л. Восс не согласился с выбором Мууса Desmoteuthis, но также ошибочно последовал за Берри и, таким образом, использовал род Megalocranchia[7]. В 1962 году бельгийский малаколог Уильям Адам не согласился с Воссом и Муусом и использовал Verilliteuthis Берри для образца, пойманного у побережья Анголы (в настоящее время идентифицированного как Teuthowenia maculata). В том же году британский теутолог Малкольм Р. Кларк также не согласился с Муусом и вместе с другими видами перенёс Teuthowenia megalops в род Стенструпа Taonius как Taonius megalops, наряду с Taonius pavo[50]. Он также признал Desmoteuthis tenera и Leachia hyperborea как синонимы под Taonius megalops. Новая комбинация Кларка была основным именем, использовавшимся последующими авторами до 1985 года, когда американский малаколог Нэнси А. Восс, наконец, прояснила запутанную таксономическую историю семейства Cranchiidae, сохранив Teuthowenia Хуна. Допустимая комбинация для вида в настоящее время — Teuthowenia megalops[2].
Примечания
- Clyde F.E. Roper; Patrizia Jereb. Family Cranchiidae // Cephalopods of the world. An annotated and illustrated catalogue of cephalopod species known to date. Volume 2: Myopsid and Oegopsid Squids (англ.) / Clyde F.E. Roper; Patrizia Jereb. — Food and Agriculture Organization of the United Nations, 2010. — Vol. Vol. 2. — P. 176—177. — (FAO Species Catalogue for Fishery Purposes No. 4). — ISBN 9789251067208.
- Nancy A. Voss. Systematics, biology and biogeography of the cranchiid cephalopod genus Teuthowenia (Oegopsida) (англ.) // Bulletin of Marine Science : journal. — 1985. — Vol. 36, no. 1. — P. 1—85.
- Edward Stuart Russell. Preliminary notice on the cephalopoda collected by the fishery cruiser Goldseeker 1903–1908 (англ.) // Annals and Magazine of Natural History : journal. — Taylor & Francis, 1909. — Vol. 3, no. 17. — P. 446—455. — doi:10.1080/00222930908692607.
- Richard E. Young & Katharina M. Mangold (1922–2003). Teuthowenia megalops. The Tree of Life Web Project. Дата обращения: 14 марта 2012.
- Michael Vecchione; Gerhard Pohle. Midwater cephalopods in the western North Atlantic Ocean off Nova Scotia (англ.) // Bulletin of Marine Science : journal. — 2002. — Vol. 71, no. 2. — P. 883—892.
- Rikke Petri Frandsen; Karsten Zumholz. Cephalopods in Greenland Waters – a field guide (англ.). — Pinngortitaleriffik, Greenland Institute of Natural Resources, 2004. — P. 17. — (Technical report no. 58). — ISBN 978-8791214103.
- Gilbert L. Voss. Bermudan cephalopods (неопр.) // Fieldiana Zoology. — 1960. — Т. 39, № 40. — С. 419—446.
- Teuthowenia megalops. Mollusca of the North Sea. Marine Species Identification Portal. Дата обращения: 13 марта 2012.
- Teuthowenia. Smithsonian National Museum of Natural History. Дата обращения: 17 марта 2012.
- Teuthowenia megalops. FishBase (23 марта 2010). Дата обращения: 16 марта 2012. (недоступная ссылка)
- Moiseev, Sergey Ivanovich. Observation of the vertical distribution and behavior of nektonic squids using manned submersibles (англ.) // Bulletin of Marine Science : journal. — 1991. — Vol. 49, no. 1—2. — P. 446—456.
- Jon A. Moore; Michael Vecchione; Karsten E. Hartel; Bruce B. Collette; John K. Galbraith; Ruth Gibbons; Mary Turnipseed. Biodiversity of Bear Seamount, New England Seamount Chain: results of exploratory trawling (англ.) // Journal of Northwest Atlantic Fishery Science : journal. — 2003. — Vol. 31. — P. 363—372. — doi:10.2960/J.v31.a28. Архивировано 3 декабря 2013 года.
- Uwe Piatkowski, Michael Vecchione; Richard E. Young. Community and species diversity of deepwater cephalopods along the northern mid-Atlantic Ridge (англ.). — International Council for the Exploration of the Sea, 2006. — Vol. D:16. — (ICES Conference and Meeting (CM) Documents).
- Cruise Report – RV G.O. Sars, June 2004 // MAR-ECO cruise to the Mid-Atlantic Ridge: Iceland–Azores (англ.). — MAR-ECO, Census of Marine Life, 2004. — (Toktrapport/Havforskningsinstituttet Number 19).
- Martin A. Collins; Cynthia Yau; Peter R. Boyle; Dagmar Friese; Uwe Piatkowski. Distribution of cephalopods from plankton surveys around the British Isles (англ.) // Bulletin of Marine Science : journal. — 2002. — Vol. 71, no. 1. — P. 239—254.
- Francis Neat; Finlay Burns; Jim Drewery. The Deepwater Ecosystem of the Continental Shelf Slope and Seamounts of the Rockall Trough: A Report on the Ecology and Biodiversity Based on FRS Scientific Surveys (англ.). — Fisheries Research Services. — (Fisheries Research Services Internal Report No 02/08).
- Colm Lordan; Steve Warnes; Tom F. Cross; Gavin M. Burnell. The distribution and abundance of cephalopod species caught during demersal trawl surveys west of Ireland and in the Celtic Sea (англ.). — 2001. — (Irish Fisheries Investigation Series No. 8). Архивированная копия (недоступная ссылка). Дата обращения: 4 августа 2019. Архивировано 20 ноября 2007 года.
- Ana Moreno; Antonina dos Santos; Uwe Piatkowski; A. Miguel P. Santos; Henrique Cabral. Distribution of cephalopod paralarvae in relation to the regional oceanography of the western Iberia (англ.) // Journal of Plankton Research : journal. — 2009. — Vol. 1. — P. 73—91. — doi:10.1093/plankt/fbn103.
- Lee C. Hastie; Graham J. Pierce; Jianjun Wang; Isabel Bruno; Ana Moreno; Uwe Piatkowski; Jean-Paul Robin. Cephalopods in the north-eastern Atlantic: species, biogeography, ecology, exploitation and conservation (англ.). — 2009. — Vol. 47. — P. 111—190. — (Oceanography and Marine Biology - An Annual Review). — ISBN 978-1-4200-9421-3. — doi:10.1201/9781420094220.ch3.
- Michael Vecchione; Odd Aksel Bergstad; Ingvar Byrkjedal; Tone Falkenhaug; Andrey V. Gebruk; Olav Rune Godø; Astthor Gislason; Mikko Heino; Age S. Høines; Gui M. M. Menezes; Uwe Piatkowski; Imants G. Priede; Henrik Skov; Henrik Søiland; Tracey Sutton; Thomas de Lange Wenneck. Biodiversity patterns and processes on the Mid-Atlantic Ridge // Life in the World's Oceans: Diversity, Distribution, and Abundance (англ.) / Alasdair McIntyre. — John Wiley and Sons, 2010. — P. 104, 111. — ISBN 9781405192972.
- Michael Vecchione; Clyde F.E. Roper. Cephalopods observed from submersibles in the western North Atlantic (англ.) // Bulletin of Marine Science : journal. — 1991. — Vol. 49, no. 1—2. — P. 433—445.
- Richard E. Young & Katharina M. Mangold (1922–2003). Teuthowenia Chun, 1910. The Tree of Life Web Project. Дата обращения: 17 марта 2012.
- Takashi Okutani; Dhugal Lindsay. Cephalopods Collected by the Submersibles and ROVs of Japan Agency for Marine-Earth Science & Technology -Annotated Catalogue up to 2008 - (англ.) // JAMSTEC Report of Research and Development : journal. — 2010. — Vol. 10. — P. 23—32. — doi:10.5918/jamstecr.10.23.
- Richard E. Young. The systematics and areal distribution of pelagic cephalopods from the seas of southern California (англ.) // Smithsonian Contributions to Zoology : journal. — 1972. — Vol. 97, no. 97. — P. 1—159. — doi:10.5479/si.00810282.97.
- Eric James Denton; John B. Gilpin-Brown; Trevor I. Shaw. A buoyancy mechanism found in cranchid squid (англ.) // Proceedings of the Royal Society : journal. — 1969. — Vol. B, no. 1036. — P. 271—279. — doi:10.1098/rspb.1969.0093.
- Richard E. Young & Katharina M. Mangold (1922–2003). Cranchiid Buoyancy. The Tree of Life Web Project. Дата обращения: 17 марта 2012.
- Cephalopods in Action. Smithsonian Institution. Дата обращения: 16 марта 2012.
- Michael Vecchione; Clyde F. E. Roper; Michael J. Sweeney; C. C. Lu. Distribution, Relative Abundance and Developmental Morphology of Paralarval Cephalopods in the Western North Atlantic Ocean (англ.). — National Oceanic and Atmospheric Administration, U.S. Department of Commerce, 2001. — P. 37—38. — (NOAA Technical Report NMFS 152). Архивированная копия (недоступная ссылка). Дата обращения: 5 августа 2019. Архивировано 9 августа 2017 года.
- Giambattista Bello. Role of cephalopods in the diet of the swordfish, Xiphias gladius, from the eastern Mediterranean Sea (англ.) // Bulletin of Marine Science : journal. — 1991. — Vol. 49, no. 1—2. — P. 312—324.
- Clarke, Malcolm R. Oceanic cephalopod distribution and species diversity in the eastern North Atlantic (англ.) // Arquipélago – Life and Marine Sciences : journal. — 2006. — No. 23A. — P. 27—46.
- Roger T. Hanlon; John B. Messenger. Cephalopod Behaviour (неопр.). — Cambridge University Press, 1998. — С. 86. — ISBN 9780521645836.
- Peter Noel Dilly. Taonius megalops, a squid that rolls up into a ball (англ.) // Nature : journal. — 1972. — Vol. 237, no. 5355. — P. 403—404. — doi:10.1038/237403a0.
- Peter Batson. Deep New Zealand: Blue Water, Black Abyss (англ.). — Canterbury University Press, 2003. — P. 120. — ISBN 9781877257094.
- Otto Mörch. Om Cranchia megalops Prosch // Videnskabelige Meddelelse den naturhistoriske Forening i Kjöbenhavn, for Aarene 1849 og 1850 (датск.). — Bianco Luno. — С. 57—64.
- Victor Prosch. Nogle nye Cephalopoder, beskrevne og anatomisk undersögte (датск.) // Det Kongelige Danske Videnskabernes Selskabs Skrifter. Naturvidenskabelig og Mathematisk Afdeling. — Т. 5, № 1. — С. 53—72.
- Steenstrup, J. J. Hectocotylus-formation in Argonauta and Tremoctopus explained by observations on similar formations in the Cephalopoda in general (англ.) // Annals and Magazine of Natural History : journal. — Taylor & Francis. — Vol. 2, no. 116. — P. 81—114. — doi:10.1080/00222935709487882.
- Steenstrup, J. J. Overblik over de i Kjöbenhavns Museer opbevarede Blaeksprutter fra det aabne Hav (1860–1861) (датск.) // Oversigt over Det Kongelige Danske Videnskabernes Selskabs Forhandlinger og Dets Medlemmers Arbeider. — С. 69—86. (недоступная ссылка)
- M.J. Imber. The squid families Cranchiidae and Gonatidae (Cephalopoda: Teuthoidea) in the New Zealand region (англ.) // New Zealand Journal of Zoology : journal. — 1978. — Vol. 5, no. 3. — P. 445—484. — doi:10.1080/03014223.1978.10428331.
- George W. Tryon. Manual of Conchology; Structural and Systematic: With Illustrations of the Species. Vol. 1. Cephalopoda (англ.). — George W. Tryon, 1879. — P. 162.
- Addison Emery Verrill. The cephalopods of the north-eastern coast of America (англ.) // Transactions of the Connecticut Academy of Arts and Sciences : journal. — 1878–1882. — Vol. 5. — P. 177—257.
- William Evans Hoyle. On Loligopsis and some other genera (неопр.) // Proceedings of the Royal Physical Society of Edinburgh. — 1883–1885. — Т. 8. — С. 313—333.
- William Evans Hoyle. Report on the cephalopoda collected by H.M.S. Challenger during the years 1873–76 // The Voyage of the H.M.S. Challenger (неопр.) / John Murray. — 1886. — С. 126—129. — doi:10.5962/bhl.title.46542.
- Nancy A. Voss. A generic revision of the Cranchidae (Cephalopoda; Oegopsida) (англ.) // Bulletin of Marine Science : journal. — 1980. — Vol. 30, no. 2. — P. 365—412.
- Carl Chun. Die Cephalopoden. I. Teil: Oegopsida. II. Teil. Myopsida, Octopoda // Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer Valdivia 1898–1899 (нем.). — Gustav Fischer, 1910. — S. 126—129. — doi:10.5962/bhl.title.13499.
- Samuel S. Berry. Some necessary changes in cephalopod nomenclature (англ.) // Science. — 1912. — Vol. 36, no. 932. — P. 643—646. — doi:10.1126/science.36.932.643. — PMID 17842183.
- Georg Johann Pfeffer. Die Cephalopoden der Plankton-Expedition. Zugleich eine monographische Übersicht der Oegopsiden Cephalopoden (нем.). — Lipsius & Tischer, 1912. — doi:10.5962/bhl.title.10474.
- Johannes Thiele. Handbook of Systematic Malacology (неопр.). — Gustav Fischer Verlag, 1934. — Т. III. — С. 1480, 1483.
- Bent J. Muus. Biodiversity patterns and processes on the Mid-Atlantic Ridge // Cephalopoda. Suborder Teuthoidea. Families: Ommastrephidae, Chiroteuthidae, Cranchiidae (англ.). — International Council for the Exploration of the Sea, 1963. — (Zooplankton Sheet 96).
- Martina A. Roeleveld. Cephalopoda from the tropical eastern Atlantic Ocean (англ.) // Galathea Report : journal. — 1977. — Vol. 14. — P. 123—132. Архивировано 6 июля 2017 года.
- Gilbert L. Voss. A New Species of Cranchiid Squid, Phasmatopsis lucifer, from the Gulf of Mexico (англ.) // Bulletin of Marine Science of the Gulf and Caribbean : journal. — 1963. — Vol. 13, no. 1. — P. 77—83.