Mad1
Mad1 — белок дрожжей и других эукариот, который участвует в прохождении контрольной точки сборки веретена деления (англ. spindle assembly checkpoint, SAC)[1]. Эта контрольная точка контролирует прикрепление микротрубочек к хромосомам и препятствует вхождению клетки в анафазу до тех пор, пока веретено деления не будет полностью собрано. Название Mad дано за то, что у клеток, мутантных по этому белку, наблюдается дефектность блокировки митоза (англ. mitotic arrest deficient, Mad) при деполимеризации микротрубочек. MAD1 рекрутирует ингибитор анафазы Mad2 к свободным кинетохорам, не соединившимся с микротрубочками веретена, и имеет важное значение для образования комплекса Mad2-Cdc20 in vivo, но не in vitro. In vivo Mad1 действует как конкурентный ингибитор комплекса Mad2-Cdc20[2]. Mad1 фосфорилируется киназой Mps1, что вместе с другими процессами приводит к формированию комплекса митотической контрольной точки (англ. mitotic checkpoint complex, MCC). Таким образом он ингибирует активность комплекса стимуляции анафазы/циклосомы (APC/C). Гомологи Mad1 эволюционно консервативны у эукариот от дрожжей до млекопитающих. Регулируя клеточный цикл и расхождение хромосом, а также выполняя некоторые интерфазные функции, Mad1 оказывается вовлечённым в развитие многих опухолей и генетических заболеваний (например, связанных с анеуплоидией)[3][4].
Функции
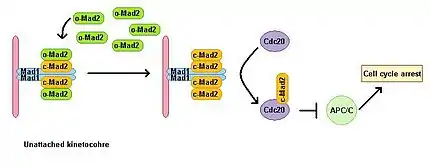
Эукариотические клетки демонстрируют блокировку митоза в присутствии ингибиторов полимеризации микротрубочек. Контрольная точка сборки веретена (SAC) следит за состоянием веретена и связывает переход от метафазы к анафазе с правильным биполярным прикреплением всех кинетохоров к митотическому веретену. SAC ингибирует активность комплекса стимуляции анафазы (APC/C), предотвращая деградацию последующих эффекторов, которые в противном случае привели бы к вступлению в анафазу и завершению митоза. Нарушение работы Mad1 приводит к потере функции SAC. Mad1 локализуется преимущественно на одиноких кинетохорах и запускает остановку митоза в случае даже всего лишь одного одинокого кинетохора. Mad1 рекрутирует важный компонент SAC — Mad2 — к одиноким кинетохорам (см. рис.) и индуцирует усиление сигнала остановки митоза. Существует пул свободного цитоплазматического Mad2 в неактивной открытой конформации, называемой о-Mad2. При связывании с Mad1 Mad2 принимает активную конформацию, называемую закрытой (c-MAD2), и образует гетеротетрамер двух Mad1 и двух c-Mad2-субъединиц. Гетеротетрамер из MAD1-c-Mad2 очень стабилен и работает как каталитический рецептор для свободного цитоплазматического о-Mad2. Свободный o-Mad2 связывается с этим рецептором и изменяет свою конформацию в активную закрытую форму. Этот второй с-MAD2 перевешивается на циклин Cdc20 по ещё не известному механизму и формирует комплекс Cdc20-с-Mad2. Этот комплекс является важнейшим компонентом митотического комплекса контрольной точки (MCC). МСС связывает и ингибирует APC/C и, следовательно, блокирует дальнейшее течение митоза[5][6].
Кроме участия в регуляции митоза, Mad1 имеет и некоторые интерфазные функции. В частности, установлено, что в интерфазе в аппарате Гольджи содержится Mad1. В отличие от цитоплазматического Mad1, Mad1 из аппарата Гольджи не зависит от Mad2. Недостаток Mad1 вызывает нарушения секреции интегрина α5 и, как следствие, нарушение клеточного прикрепления и адгезии и уменьшение подвижности клеток. Напротив, его сверхэкспрессия приводит к усилению направленной миграции клеток[3].
Было показано, что у растений Mad1 участвует в эндополиплоидизации и цветении путём взаимодействия с белком Mos1 — отрицательным регулятором растительного иммунитета[7].
Регуляция
Существуют две киназы контрольной точки, вовлеченные в регуляцию функционирования Mad1 посредством фосфорилирования[8]. Одна из них, Mps1, фосфорилирует MAD1 как in vitro, так и in vivo и, как полагают, регулирует локализацию Mad1 и Mad2 на кинетохорах и динамику их взаимодействия. Другая киназа, BUB1, рекрутирует Mad1 к кинетохорам и активирует его, если есть неприкреплённые кинетохоры[5]. Если кинетохор прикреплен к веретену, ингибитор SAC p31comet ингибирует конформационную перестройку Mad1 в Mad2 и предотвращает связывание Mad2 с Cdc20[5].
Структурные особенности и механизм
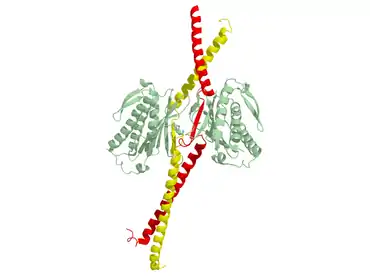
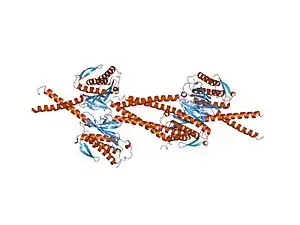
Биохимическими методами 1995 году было предсказано, что белок Mad1 представляет собой двуспиральный белок с характерной формы стержня, массой 90 кДа и включающий 718 аминокислотных остатков[9][1] . Затем в 2002 году была опубликована кристаллическая структура человеческих Mad1 в комплексе с Mad2, образующими тетрамер (см. рис.). В связи с экспериментальными ограничениями эта структура показывает только с 484-го по 584-й аминокислотные остатки, входящие в состав Mad1. Удлиненные мономеры MAD1 плотно скрепляются параллельной двойной спиралью с участием N-концевых альфа-спиралей. Цепи MAD1 указывают от двойных спиралей к их лигандам Mad2, образуя два субкомплекса с Mad2. Отрезок между альфа-спиралями 1 и 2 содержит домен для связывания Mad2. Первая часть этого связывающего домена является гибкой и принимает разные конформации, обусловливающие асимметрию комплекса. С использованием термодинамических исследований было показано, что Mad1 способен замедлять скорости образования комплекса Mad2-Cdc20 и, следовательно, выступает в качестве конкурентного ингибитора in vivo. Более того, установлено, что сайты связывания Mad1 и Mad2 находятся внутри комплекса, возможно, делая сайты связывания с Cdc20 недоступными для него. Связывание Mad1-Mad2 необычно тем, что C-конец Mad2 покрывает Mad1. В связи с этим предполагается, что неактивный комплекс Mad1-Mad2 не способен высвобождать Mad2, следовательно, для этого необходим новый, пока плохо изученный механизм конформационных перестроек комплекса[2].
Примечания
- Hardwick K. G., Murray A. W. Mad1p, a phosphoprotein component of the spindle assembly checkpoint in budding yeast. (англ.) // The Journal of cell biology. — 1995. — Vol. 131, no. 3. — P. 709—720. — PMID 7593191.
- Sironi L., Mapelli M., Knapp S., De Antoni A., Jeang K. T., Musacchio A. Crystal structure of the tetrameric Mad1-Mad2 core complex: implications of a 'safety belt' binding mechanism for the spindle checkpoint. (англ.) // The EMBO journal. — 2002. — Vol. 21, no. 10. — P. 2496—2506. — doi:10.1093/emboj/21.10.2496. — PMID 12006501.
- Wan J., Zhu F., Zasadil L. M., Yu J., Wang L., Johnson A., Berthier E., Beebe D. J., Audhya A., Weaver B. A. A Golgi-localized pool of the mitotic checkpoint component Mad1 controls integrin secretion and cell migration. (англ.) // Current biology : CB. — 2014. — Vol. 24, no. 22. — P. 2687—2692. — doi:10.1016/j.cub.2014.09.052. — PMID 25447996.
- Avram S., Mernea M., Mihailescu D. F., Seiman C. D., Seiman D. D., Putz M. V. Mitotic checkpoint proteins Mad1 and Mad2 - structural and functional relationship with implication in genetic diseases. (англ.) // Current computer-aided drug design. — 2014. — Vol. 10, no. 2. — P. 168—181. — PMID 24724894.
- Musacchio A., Salmon E. D. The spindle-assembly checkpoint in space and time. (англ.) // Nature reviews. Molecular cell biology. — 2007. — Vol. 8, no. 5. — P. 379—393. — doi:10.1038/nrm2163. — PMID 17426725.
- Yu H. Structural activation of Mad2 in the mitotic spindle checkpoint: the two-state Mad2 model versus the Mad2 template model. (англ.) // The Journal of cell biology. — 2006. — Vol. 173, no. 2. — P. 153—157. — doi:10.1083/jcb.200601172. — PMID 16636141.
- Bao Z., Zhang N., Hua J. Endopolyploidization and flowering time are antagonistically regulated by checkpoint component MAD1 and immunity modulator MOS1. (англ.) // Nature communications. — 2014. — Vol. 5. — P. 5628. — doi:10.1038/ncomms6628. — PMID 25429892.
- Bharadwaj R., Yu H. The spindle checkpoint, aneuploidy, and cancer. (англ.) // Oncogene. — 2004. — Vol. 23, no. 11. — P. 2016—2027. — doi:10.1038/sj.onc.1207374. — PMID 15021889.
- Chen R. H., Shevchenko A., Mann M., Murray A. W. Spindle checkpoint protein Xmad1 recruits Xmad2 to unattached kinetochores. (англ.) // The Journal of cell biology. — 1998. — Vol. 143, no. 2. — P. 283—295. — PMID 9786942.
Литература
- Yang C., Hao J., Kong D., Cui X., Zhang W., Wang H., Guo X., Ma S., Liu X., Pu P., Xu B. ATM-mediated Mad1 Serine 214 phosphorylation regulates Mad1 dimerization and the spindle assembly checkpoint. (англ.) // Carcinogenesis. — 2014. — Vol. 35, no. 9. — P. 2007—2013. — doi:10.1093/carcin/bgu087. — PMID 24728176.
- Kuijt T. E., Omerzu M., Saurin A. T., Kops G. J. Conditional targeting of MAD1 to kinetochores is sufficient to reactivate the spindle assembly checkpoint in metaphase. (англ.) // Chromosoma. — 2014. — Vol. 123, no. 5. — P. 471—480. — doi:10.1007/s00412-014-0458-9. — PMID 24695965.
- Ballister E. R., Riegman M., Lampson M. A. Recruitment of Mad1 to metaphase kinetochores is sufficient to reactivate the mitotic checkpoint. (англ.) // The Journal of cell biology. — 2014. — Vol. 204, no. 6. — P. 901—908. — doi:10.1083/jcb.201311113. — PMID 24637323.