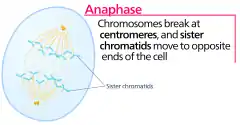
Анафаза
Анафа́за — фаза митотического деления эукариотических клеток, в которой сестринские хроматиды синхронно расходятся с образованием двух дочерних хромосом, которые неспешно растаскиваются к противоположным полюсам веретена деления. В ходе анафазы кинетохорные микротрубочки укорачиваются, а полюса удаляются друг от друга, таким образом, оба процесса вносят свой вклад в расхождение хроматид[1].
Анафаза
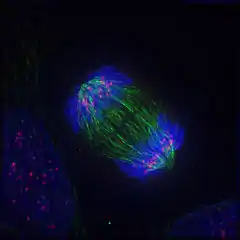
Описание
Начало расхождения сестринских хроматид знаменует собой переход от метафазы к анафазе. Анафаза — самая короткая фаза митоза. Хромосомы движутся равномерно, со скоростью 0,5—2 мкм в секунду. Движущиеся хромосомы часто принимают V-образную форму: центромера смотрит в сторону полюсов деления, а плечи как бы откинуты к центру веретена. Если по каким-то причинам плечо хромосомы претерпело разрыв, то она не будет участвовать в движении и останется в центральной зоне. У некоторых высших растений, например, у ожики, выраженной центромеры нет, и микротрубочки веретена взаимодействуют со многими точками поверхности хромосом. В этом случае хромосомы лежат поперёк волокон веретена[2].
Комплекс стимуляции анафазы (APC/C) запускает расхождение, убиквитинилируя и инициируя разрушение нескольких белков-регуляторов митоза. Если во время метафазы белки когезины удерживают сестринские хроматиды вместе и противодействуют силам, тянущим их к полюсам, то в начале анафазы наблюдается резкое нарушение их когезии, позволяющее им разъединиться и начать расходиться к противоположным полюсам клетки. Уход когезина из сестринских хроматид запускается сепаразой, которая до анафазы связана с секурином и потому неактивна. APC/C запускает деградацию секурина и, следовательно, расхождение хроматид. APC/C также запускает разрушение S- и M-циклинов, в результате чего циклинзависимые киназы (Cdk) в анафазе почти неактивны. Из-за этого фосфатазы дефосфорилируют белки-мишени Cdk, что необходимо для окончания митоза. Активация APC/C происходит под действием белка Cdc20, который связывается с APC/C в митозе и активирует его. В ходе этого процесса сначала увеличивается экспрессия Cdc20, потом фосфорилирование APC/C M-Cdk и другими киназами способствует его связыванию с Cdc20 с образованием активного комплекса[3].
В основе расхождения сестринских хроматид лежат два независимых перекрывающихся процессов: анафазы А и анафазы В. Анафаза А — это направленное к полюсам движение хромосом, сопровождающееся укорочением микротрубочек кинетохора. Движение хромосом в анафазе А зависит от двух сил, направленных к полюсам: силы деполимеризации микротрубочек в кинетохоре и силы движения микротрубочек к полюсу веретена деления, где происходит деполимеризация минус-концов (микротрубочковый ток). Для движения хромосом в анафазе А необходимо присутствие АТФ и достаточная концентрация ионов Ca2+[4]. В эмбриональных клетках расхождение хромосом в анафазе А происходит за счёт микротрубочкового тока, а в дрожжах и соматических клетках позвоночных — от кинетохорных сил. Анафаза В включает расхождение самих полюсов веретена деления после разделения сестринских хроматид и расхождения дочерних хромосом на некоторое расстояние. Отталкивание полюсов друг от друга обеспечивается работой моторных белков кинезинов-5, движущихся к плюс-концам микротрубочек. Динеины, связывающие астральные микротрубочки с клеточным кортексом, также вносят свой вклад в расхождение хромосом. В анафазе В расстояние между двумя группами хроматид становится ещё больше, это необходимо для формирования борозды деления[5]. Относительные вклады анафазы А и анафазы В в расхождение хромосом варьируют в клетках разных типов. Так, в клетках млекопитающих анафаза В начинается вскоре после анафазы А и завершается при двукратном увеличении размеров веретена по сравнению с таковой в метафазе. В то же время в клетках дрожжей и других одноклеточных организмов для разделения хромосом в анафазе используется преимущественно анафаза В, и веретено деления в анафазе может увеличиться в 15 раз по сравнению с метафазой[6]. В клетках растений анафазы В нет[7].
Примечания
- Альбертс и др., 2013, с. 1649.
- Ченцов, 2005, с. 440—441.
- Альбертс и др., 2013, с. 1670—1671.
- Ченцов, 2005, с. 441.
- Кассимерис, Лингаппа, Плоппер, 2016, с. 680.
- Альбертс и др., 2013, с. 1673—1674.
- Ченцов, 2005, с. 442.
Литература
- Кассимерис Л., Лингаппа В. Р., Плоппер Д. . Клетки по Льюину. — М.: Лаборатория знаний, 2016. — 1056 с. — ISBN 978-5-906828-23-1.
- Альбертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. Молекулярная биология клетки: в 3-х томах. Т. II. — М.: Ижевск: НИЦ «Регулярная и хаотическая динамика», Институт компьютерных исследований, 2013. — 992 с. — ISBN 978-5-4344-0113-5.
- Ченцов Ю. С. Введение в клеточную биологию. — М.: ИКЦ «Академкнига», 2005. — 495 с. — ISBN 5-94628-105-4.
| Фазы |
|  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Регуляторы |
| ||||||||||