Эволюционная теория пола В. А. Геодакяна
Эволюционная теория пола была предложена Вигеном Геодакяном в 1965 году. Теория объясняет[1] с единых позиций многие явления, связанные с полом: половой диморфизм в норме[2] и патологии[3][4], соотношение полов[5], дифференциальную смертность и норму реакции полов[6], роль половых хромосом[7] и половых гормонов, асимметрию мозга[8] и рук[9], отцовский эффект|реципрокные эффекты и психологические и социальные различия между полами[10][11].
История вопроса
С 1965 года было опубликовано более 150 работ, посвященных теории пола и смежным вопросам — длительности жизни, дифференциации мозга и рук, половым хромосомам, механизмам регуляции у растений и животных, порокам сердца и другим болезням и даже культуре, были сделаны сообщения на многих отечественных и международных конгрессах, конференциях и симпозиумах[12][13]. Две конференции были посвящены исключительно теории (Санкт-Петербург, Россия, 1990, 1992)[14]. Теория была включена в учебники[15][16][17][18][19][20], учебные пособия[21][22][23][24][25][19] и программы преподавания ряда университетов и институтов[26][27]. О теории неоднократно писали на страницах периодической печати[12]. На телевидении были показаны 3 интервью в программе А. Гордона[28][29][30].
Теория цитируется во многих монографиях[31][32][33][34][35][36] и используется в различных областях научной деятельности: эволюции полового размножения[31][37], биологии растений[38] и животных[39], медицине[40][41], социальной психологии[42][43][44][45][46][47], педагогике[48] и др.
Принцип сопряжённых подсистем
Теория основана на принципе сопряжённых подсистем, которые эволюционируют асинхронно. Мужской пол является оперативной подсистемой популяции, женский пол — консервативной подсистемой. Новая информация от среды попадает сначала к мужскому полу и только после многих поколений передается женскому, поэтому эволюция мужского пола предшествует эволюции женского. Этот сдвиг во времени (две фазы эволюции признака) создает две формы признака (мужскую и женскую) — половой диморфизм в популяции. Эволюционная «дистанция» между подсистемами необходима для поиска и проверки новаций.
Интерпретация полового диморфизма как филогенетической «дистанции» между полами, как эволюционные «новости», которые уже попали в мужскую подсистему, но ещё не переданы женской, применимо для всех признаков растений, животных и человека по которым наблюдается половой диморфизм. Только в случае видовых признаков закономерность проявляется в области патологии, популяционных — в норме, а по половым признакам — в виде «отцовского эффекта».
Теория связывает основные характеристики раздельнополой популяции: соотношение полов, дисперсию полов и половой диморфизм, с условиями среды и эволюционной пластичностью популяции. В оптимальных, стабильных условиях среды эти характеристики минимальны, то есть падает рождаемость (одновременно и смертность) мальчиков, сокращается их разнообразие и разница между мужским и женским полом. Все это снижает эволюционную пластичность популяции. В экстремальных условиях, когда для быстрой адаптации требуется высокая эволюционная пластичность, идут обратные процессы: растут одновременно рождаемость и смертность (то есть «оборачиваемость») мужского пола, его разнообразие, чётче становится половой диморфизм.
Анализ проблемы пола
Понятие пола включает два фундаментальных явления: половой процесс (слияние генетической информации двух особей) и половую дифференциацию (разделение этой информации на две части). В зависимости от присутствия или отсутствия этих явлений, множество существующих способов размножения можно разделить на три основные формы: бесполое, гермафродитное и раздельнополое. Половой процесс и половая дифференциация явления разные и, по своей сути, диаметрально противоположные. Половой процесс создает разнообразие генотипов, и в этом состоит признаваемое многими учёными преимущество половых способов перед бесполыми. Половая дифференциация, наложив запрет на однополые комбинации (мм, жж), наоборот, снижает его вдвое (явление известное в англоязычной литературе как «two-fold cost of sex»). То есть, при переходе от гермафродитного размножения к раздельнополому утрачивается минимум половина разнообразия.
Тогда, непонятно, что дает разделение на два пола, если оно ухудшает вдвое основное достижение полового размножения? Почему все прогрессивные в эволюционном плане виды животных: (млекопитающие, птицы, насекомые) и растений (двудомные) — раздельнополы, в то время, как явные преимущества количественной эффективности и простоты у бесполых форм, а разнообразия потомства — у гермафродитных?
Чтобы решить загадку раздельнополости необходимо объяснить, что дает дифференциация, а для этого необходимо понять преимущества раздельнополости перед гермафродитизмом. Это значит, что раздельнополость, которую тщетно пытаются понять как лучший способ размножения, вовсе таковым не является. Это — эффективный способ эволюции[49][50].
Консервативно-оперативная специализация полов
Разделение на два пола это специализация по сохранению и изменению информации в популяции. Один пол должен быть информационно более тесно связан со средой, и быть более чувствительным к её изменениям. Повышенная смертность мужского пола от всех факторов среды позволяет считать его оперативной экологической подсистемой популяции. Женский пол, как более стабильный, является консервативной подсистемой и сохраняет существующее распределение генотипов в популяции.
В эволюции пола на разных стадиях и уровнях организации появился целый ряд механизмов, которые последовательно обеспечивали более тесную связь женского пола с генеративным (консервативным) потоком, а мужского — с экологическим (оперативным). Так, у мужского пола, по сравнению с женским, выше частота мутаций, меньше аддитивность наследования родительских признаков, у́же норма реакции, выше агрессивность и любознательность, активнее поисковое, рискованное поведение и другие качества, «приближающие к среде». Все они, целенаправленно вынося мужской пол на периферию распределения, обеспечивают ему преимущественное получение экологической информации.
Другая группа особенностей — огромная избыточность мужских гамет, их малые размеры и высокая подвижность, большая активность и мобильность самцов, их склонность к полигамии и другие этолого-психологические свойства[11]. Длительные периоды беременности, кормления и заботы о потомстве у самок, фактически повышая эффективную концентрацию мужских особей, превращают мужской пол в «избыточный», стало быть, «дешевый», а женский — в дефицитный и более ценный.
В результате консервативно-оперативной специализации полов происходит их асинхронная эволюция: новые признаки появляются сначала в оперативной подсистеме (мужской пол) и только потом попадают в консервативную (женский пол).
Мужской пол остается в опасных зонах и подвергается действию отбора. После действия отбора уменьшается доля мужских особей и сужается их генотипическая дисперсия. В движущей среде преобразования затрагивают и дисперсии полов и средние значения признака: норма реакции создает временный, фенотипический половой диморфизм, отбор — генотипический. Мужской пол получает новую экологическую информацию. Повышение смертности мужского пола увеличивает рождаемость мужских особей за счет отрицательной обратной связи.
Эволюционная роль половых хромосом и половых гормонов
Половой процесс и половая дифференциация действуют в противоположных направлениях: первый увеличивает разнообразие генотипов, а вторая — минимум в два раза ухудшает его. Поэтому называть «неодинаковую» пару гомологичных хромосом (ХY, ZW) «половыми», только из-за того, что они определяют пол, не совсем корректно. Гораздо больше оснований считать их «антиполовыми», так как именно они ухудшают главное достижение пола — комбинаторику признаков. Основная роль половых хромосом — эволюционная,— создание двух сдвинутых по времени форм (женской и мужской), для экономной эволюции.
Пол зиготы определяют при зачатии половые хромосомы. Дальше, до конца онтогенеза, полом управляют половые гормоны. У млекопитающих базовый пол — гомогаметный (XX) — женский; а производный пол — гетерогаметный (XY) — мужской. Он запускается Y-хромосомой, превращающей «бесполые» зачатки гонад эмбриона в семенники, продуцирующие андрогены. При отсутствии Y-хромосомы, те же ткани превращаются в яичники, продуцирующие эстрогены. У птиц базовый пол тоже гомогаметный (ZZ), но — мужской; а производный женский пол имеет гетерогаметную конституцию (ZW). Он запускается W-хромосомой, превращающей зачатки в яичники, продуцирующие эстрогены. При отсутствии W-хромосомы, те же ткани превращаются в семенники, продуцирующие андрогены. То есть, у млекопитающих андрогены сдвигают самцов от самок к среде, а у птиц — эстрогены удаляют самок от самцов и среды. В обоих случаях, мужской пол является «средовым», а женский — «системным».
Половые гормоны определяют развитие не только признаков половой дифференциации (половой диморфизм), но и асимметрии мозга, рук и других частей тела (латеральный диморфизм). Эстрогены, расширяя норму реакции, позволяют женским фенотипам покинуть зоны отбора и сохраниться. Они действуют «центростремительно», удаляя и изолируя систему от среды. Андрогены — их химические антагонисты, действуют, наоборот — «центробежно», приближая систему к среде, подвергая её более интенсивному действию отбора и ускоряя эволюцию. Следовательно, андроген-эстрогенное соотношение регулирует интенсивность информационного контакта системы со средой.
Более широкая норма реакции женского пола
Более широкая норма реакции женского пола была теоретически предсказана в 1973 году[6][51]. Согласно гипотезе, у особей мужского пола доля «наследственной компоненты» должна быть больше, а «средовой» — меньше, чем у особей женского пола. Поэтому, фенотипическая дисперсия мужского пола лучше отражает его генетическое распределение. Влияние среды в онтогенезе сильнее на женский пол, поэтому любое обучение или тренировка более эффективно.
Если сравнить внутрипарную и межпарную дисперсию у однояйцевых (монозиготных) близнецов мужского и женского пола, то внутрипарная должна быть больше у женских пар, а межпарная — у мужских. Далее, фенотипическая дисперсия в чистой линии может быть относительно шире у женского пола, а в полиморфной (дикой) популяции она шире у мужского.
Два исследования, проведенные на 44 парах однояйцевых близнецов[52] и 53 однояйцевых и 38 двуяйцевых близнецов[53], подтвердили предсказание теории.
Получение экологической информации от среды
Во-первых, изменение факторов среды может элиминировать самую чувствительную к данному фактору часть особей популяции, в результате естественного отбора. Bo-вторых, изменение факторов среды, создав дискомфортные условия, может полностью или частично отстранить от размножения другую часть популяции — за счет полового отбора. В-третьих, изменившаяся среда модифицирует выжившую часть популяции, создавая морфо-физиологические, поведенческие и другие ненаследуемые адаптации, — за счет нормы реакции. Например, в холоде у животных укорачиваются хвосты, гуще становится мех, утолщается подкожная жировая прослойка. Человек использует пещеры, одежду, огонь.
Первые два процесса (элиминация и дискриминация) убирают какие-то генотипы из пула воспроизводства. Третий процесс (модификация), наоборот, позволяет каким-то генотипам сохраниться под прикрытием модифицированного фенотипа и попасть в генофонд потомства. То есть, кого-то приходится ломать, убивать, отстранять, а кого-то — гнуть, «воспитывать», переделывать.
Чтобы получить экологическую информацию от среды, мужской пол должен иметь бóльшую фенотипическую дисперсию, которая может быть следствием широкой генотипической дисперсии. Она может быть также следствием более широкой наследственной нормы реакции женских особей, которая позволяет им покинуть зоны элиминации и дискомфорта. Более широкая генотипическая дисперсия мужского пола может быть результатом более высокого уровня мутаций у мужских особей, а также того, что женские потомки наследуют родительские признаки более аддитивно.
Механизмы регуляции параметров популяции
Два механизма контролируют параметры популяции у животных — стресс и половые гормоны. Растения получают экологическую информацию от среды через количество пыльцы[54]. Конкретная природа фактора среды, по которому организм испытывает дискомфорт, для запуска этих механизмов не имеет, видимо, никакого значения, то есть безразлично, чем вызван дискомфорт — морозом, засухой, голодом или врагами. Во всех неблагоприятных условиях, при определенной интенсивности дискомфорта, развивается стрессовое состояние, то есть такая «обобщённая» экологическая информация как бы «одномерна» — только «хорошо» или «плохо».
Соотношение полов
Повышенная смертность мужского пола
В ходе онтогенеза соотношение полов у многих видов растений, животных и человека понижается. Это связано с повышенной смертностью и повреждаемостью мужских систем, по сравнению с соответствующими женскими. Эта картина наблюдается почти на всех стадиях онтогенеза и на всех уровнях организации, исследуем ли мы различные виды (человек, животные или растения), разные уровни организации (особь, орган, ткань или клетка), или устойчивость к разным вредным факторам среды (низкие и высокие температуры, голод, яды, паразиты, болезни и др.).
Гамильтон (1948) приводит обзор дифференциальной смертности полов для 70 видов, включая такие различные формы жизни, как нематоды, моллюски, ракообразные, насекомые, паукообразные, птицы, рептилии, рыбы, млекопитающие. Согласно этим данным, у 62 видов (89 %) средняя длительность жизни самцов короче, чем у самок; у большинства остальных нет разницы, и лишь в отдельных случаях длительность жизни самцов больше, чем у самок[55].
Эволюционная теория пола рассматривает повышенную смертность мужского пола, как выгодную для популяции форму информационного контакта со средой, осуществляемую через элиминацию вредным фактором среды части особей популяции. Например, все «новые» болезни, болезни «века» или «цивилизации» (инфаркт, атеросклероз, гипертония и др.[56]), как правило, болезни мужского пола.
«Оборачиваемость» мужских особей в экстремальных условиях среды
В изменчивых, экстремальных условиях среды, повышается смертность мужского пола и падает третичное соотношение полов популяции. Чем изменчивее среда, тем меньше остается в популяции мужских особей и, одновременно, тем больше их требуется для приспособления. Компенсировать понижение третичного соотношения полов можно только повысив вторичное. Иными словами, в экстремальных условиях среды будет одновременно повышаться и смертность и рождаемость мужских особей, — то есть будет расти их «оборачиваемость».
Регуляция соотношения полов популяции
В 1965 году было предположено, что наряду с прямой связью между вторичным и третичным соотношением полов существует также и регуляторная отрицательная обратная связь.
Организменные механизмы регуляции соотношения полов
Отрицательная обратная связь реализуется у растений через количество пыльцы, а у животных — через интенсивность половой деятельности, старение, сродство и гибель гамет. При этом, малое количество пыльцы, интенсивная половая деятельность самцов, свежая сперма и старые яйцеклетки должны приводить к увеличению рождаемости мужских особей[5][54].
Популяционные механизмы регуляции соотношения полов
Для реализации популяционного механизма необходимо, чтобы вероятность иметь потомка данного пола различалась у разных особей и определялась их генотипом. При этом, должна существовать обратная зависимость между репродуктивным рангом данной особи и полом её потомства: чем выше репродуктивный ранг, тем больше должно быть потомков противоположного пола. В этом случае, регуляция может осуществляться на популяционном уровне — большим или меньшим участием в размножении особей, дающих в потомстве избыток самцов или самок.
«Сечение» канала передачи информации потомству
Каждому потомку отец и мать передают примерно одинаковое количество генетической информации, но количество потомства, которому может передать генетическую информацию самец, несравненно больше количества, которому может передать информацию самка. Каждый самец, в принципе, может передать информацию всему потомству популяции, в то время, как самки такой возможности лишены. То есть, пропускная способность — «сечение» — канала связи самца с потомством значительно больше, чем сечение канала связи самки.
«Сечение» канала связи и репродуктивная структура популяции
В строго моногамной популяции число отцов и матерей равно, то есть самцы и самки имеют одинаковое «сечение канала» связи с потомством. В случае полигинии, когда отцов меньше, чем матерей, самцы имеют большее «сечение» канала связи. В случае полиандрии — все наоборот.
Онтогенетическая и филогенетическая пластичность
Широкая норма реакции делает женский пол более изменчивым и пластичным в онтогенезе. Она позволяет особям женского пола покинуть зоны элиминации и дискомфорта, собраться в зоне комфорта и уменьшить фенотипическую дисперсию и смертность.
Более узкая норма реакции мужского пола не позволяет ему уменьшить фенотипическую дисперсию. Мужские особи остаются в зонах элиминации и дискомфорта и погибают, или не оставляют потомство. Это позволяет популяции за новую информацию «платить» в первую очередь жертвой мужских особей.
Высокая онтогенетическая пластичность женского пола обеспечивает ему высокую стабильность в филогенезе. В ряду поколений, женский пол более полно сохраняет имеющееся в популяции распределение генотипов. Генотипическое распределение мужского пола меняется гораздо сильнее. Следовательно, в филогенетическом плане более изменчив, пластичен мужской пол, а в онтогенетическом — наоборот, более пластичен и изменчив женский. Такое, на первый взгляд парадоксальное, распределение ролей в филогенезе и онтогенезе, на самом деле, последовательно и непротиворечиво реализует идею специализации полов по консервативной и оперативной задачам эволюции.
Половой диморфизм
Стабильные условия среды
В стабильной среде все преобразования генетической информации затрагивают дисперсии полов, но не затрагивают средних значений признаков. Поэтому, половой диморфизм отсутствует. Возникает только разница в дисперсии, которая при переходе к следующему поколению исчезает. Однако, необходимо, чтобы генотипический половой диморфизм по норме реакции существовал заранее (в стабильной фазе), причём, генетическая информация о широкой норме реакции должна передаваться только по женской линии, а об узкой — только по мужской.
Изменяющаяся среда
В движущей среде, фенотипическое распределение мужского пола, до действия отбора, примерно повторяет исходное генотипическое распределение. Широкая норма реакции женского пола приводит к сдвигу распределения фенотипов и к появлению временного — фенотипического — полового диморфизма. Женский пол покидает зоны отбора и дискомфорта, и сохраняет спектр прошлых генотипов.
Возникшая разница между мужскими и женскими гаметами частично сохраняется и после оплодотворения, так как информация передаваемая через Y-хромосому никогда не попадает от отца к дочери[57]. В пользу того, что часть генетической информации остается в мужской подсистеме и не попадает в женскую, свидетельствует также существование реципрокных эффектов, — то, что при гибридизации не безразлично из какой породы отец, а из какой мать.
Итак, разные сечение канала и норма реакции мужского и женского пола, в движущей среде, неизбежно приводят, уже в одном поколении, к возникновению генотипического полового диморфизма. В последующих поколениях, в движущей среде, он может накапливаться и расти.
Половой диморфизм в филогенезе
Если перейти к филогенетическому масштабу времени, то у раздельнополых форм, после смены стабилизирующей среды на движущую, в течение многих поколений признак меняется только у мужского пола. У женского пола — сохраняется старое значение признака. Траектория эволюции признака раздваивается на мужскую и женскую ветви, происходит «расхождение» признака у двух полов — появление и рост генотипического полового диморфизма. Это — дивергентная фаза, в которой скорость эволюции признака больше у мужского пола.
Через некоторое время, когда исчерпываются возможности нормы реакции и других механизмов защиты женского пола, признак начинает меняться и у него. Генотипический половой диморфизм, достигнув своего оптимума, остается постоянным. Это — стационарная фаза, когда скорости эволюции признака у мужского и женского пола равны. Когда у мужского пола признак достигает нового эволюционно-стабильного значения, у женского пола он продолжает ещё меняться. Это — конвергентная фаза эволюции признака, когда скорость её больше у женского пола. Генотипический половой диморфизм постепенно уменьшается и, при слиянии признаков у двух полов, исчезает. Стало быть, фазы эволюции признака у мужского и женского пола сдвинуты во времени: у мужского пола они начинаются и кончаются раньше, чем у женского.
Так как эволюция признака всегда начинается с расширения его генотипической дисперсии и кончается её сужением, то в дивергентной фазе дисперсия шире у мужского пола, а в конвергентной — у женского. Значит, по половому диморфизму и дисперсии полов можно судить о направлении и фазе эволюции признака.
Половой диморфизм по признакам
Все признаки можно разделить на три группы по степени различия между полами.
Признаки, одинаковые у обоих полов
К первой группе отнесем те признаки, по которым между мужским и женским полом нет никакой разницы. К ним относятся качественные признаки, которые проявляются на уровне вида — общий для обоих полов план и принципиальное строение тела, число органов и многие другие. Половой диморфизм по этим признакам в норме отсутствует. Но он наблюдается в области патологии. У девочек чаще проявляются атавистические аномалии (возвраты или остановки развития), а у мальчиков — футуристические (поиск новых путей). Например, среди 4000 новорожденных детей с тремя почками девочек было в 2.5 раза больше, чем мальчиков, а среди 2000 детей с одной почкой было примерно в 2 раза больше мальчиков. Напомним, что у наших далеких предков в каждом сегменте тела имелась пара выделительных органов — метанефридиев. Следовательно, три почки у девочек — это возврат к предковому типу (атавистическое направление), а одна почка у мальчиков — футуристическая тенденция. Такая же картина наблюдается среди детей со сверхнормативным числом ребер, позвонков, зубов и т. д., то есть органов, претерпевших в процессе эволюции уменьшение числа — среди них больше девочек. Среди же новорожденных с их нехваткой больше мальчиков. Сходная картина наблюдается и в распределении врожденных пороков сердца и магистральных сосудов.
Признаки, присущие только одному полу
Ко второй группе относятся признаки, встречающиеся только у одного пола. Это первичные и вторичные половые признаки: половые органы, молочные железы, борода у человека, грива у льва, а также многие хозяйственные признаки (продукция молока, яиц, икры и т. д.). Половой диморфизм по ним носит генотипический характер, поскольку в фенотипе одного пола эти признаки отсутствуют, но наследственная информация об этих признаках записана в генотипе обоих полов. Поэтому, если они эволюционируют, то по ним должен существовать генотипический половой диморфизм. Обнаруживается он в виде реципрокных эффектов.
Признаки, присутствующие у обоих полов
Третья группа признаков находится посередине между первой (половой диморфизм отсутствует) и второй группой (половой диморфизм носит абсолютный характер). К ней относятся признаки, которые встречаются и у мужского пола, и у женского, но распределены в популяции с разной частотой и степенью выраженности. Это количественные признаки: рост, вес, размеры и пропорции, многие морфофизиологические и этолого-психологические признаки[11]. Половой диморфизм по ним проявляется как отношение их средний значений. Он справедлив для всей популяции, но может иметь обратное значение для отдельной пары особей. Именно этот половой диморфизм служит «компасом» эволюции признака. Половой диморфизм по этим признакам (например, весу) может проявляться на самых ранних этапах онтогенеза[58].
Половой диморфизм и эволюция признаков
Половой диморфизм тесно связан с эволюцией признака: он должен отсутствовать или быть минимальным для стабильных признаков и максимальным, наиболее четко выраженным, по филогенетически молодым (эволюционирующим) признакам. Как и две другие основные характеристики раздельнополой популяции — дисперсия и соотношение полов, половой диморфизм рассматривается не как константа, свойственная данному виду, как считалось раньше, а как переменная и регулируемая величина, тесно связанная с условиями среды и определяющая, в свою очередь, эволюционную пластичность признака. Поскольку в изменчивой, экстремальной среде требуется большая пластичность, чем в стабильной (оптимальной), то и половой диморфизм в стабильной среде должен уменьшаться, а в изменчивой — расти.
Половой диморфизм и репродуктивная структура популяции
Половой диморфизм должен быть связан с репродуктивной структурой популяции: у строгих моногамов он должен быть минимальным, поскольку моногамы используют специализацию полов только на уровне организма; у полигамных же видов, полнее использующих преимущества дифференциации, он должен возрастать с ростом степени полигамии.
Половой диморфизм у реципрокных гибридов («Отцовский эффект»)
По признакам, присущим только одному полу (первичные и вторичные половые признаки, а также многие хозяйственно-ценные признаки: продукция яиц, молока, икры), половой диморфизм имеет абсолютный, организменный, характер. Поскольку в фенотипе одного пола эти признаки отсутствуют, о генотипическом половом диморфизме по ним можно судить по реципрокным эффектам. Если по «старым» (стабильным) признакам генетический вклад отца в потомка в среднем несколько меньше вклада матери (из-за материнского эффекта, обусловленного цитоплазматической наследственностью, гомогаметной конституцией и утробным развитием у млекопитающих), то по «новым» признакам, согласно эволюционной теории пола, должно существовать некоторое доминирование отцовских признаков над материнскими.
Отцовский эффект установлен по алкоголизму у человека, по инстинкту насиживания, скороспелости, яйценоскости и живому весу у кур, по динамике роста, числу позвонков и длине тонкого кишечника у свиней, по удою молока и продукции молочного жира у крупного рогатого скота. Наличие отцовского эффекта по удою и яйценоскости означает не что иное, как более высокую генотипическую «удойность» у быков и «яйценоскость» у петухов, чем у коров и кур тех же пород[59].
Половой диморфизм в психологии и этологии
Так как женский пол специализируется больше по генетическому потоку информации и связям внутри популяции, у них должны быть лучше развиты язык и вербальные способности. Этологические особенности женского пола направлены на сохранение старого, уже освоенного и совершенствование уже найденных решений. Самки больше стремятся приспособиться к среде, выжить и оставить потомство. Поэтому они более податливы, больше подвержены влиянию среды и более эффективно обучаются.
С другой стороны, экологическая специализация мужского пола может объяснить их лучше развитые пространственно-зрительные способности, больше связанные со средой (защита, охота, борьба с врагами). Поведенческие особенности мужского пола направлены на изменение старого и имеют характер поиска новых решений. Они охотнее проявляют более рискованное, «исследовательское» поведение, хуже обучаются и менее конформны.
На основе эволюционного подхода был проведен анализ психологических различий между полами в вербальных и физических способностях, импульсивности и поиска ощущений[11], а также в процессе обучения[60], психологии творчества[42], различии в статусных предпочтениях[43] и стремлении к власти и контролю[61][11]. Трофимова предложила добавление к теории Геодакяна в форме понятия «пропалывание ненужностей»(redundancy prunning)[11]. Это понятие описывает тенденцию мужской части пола сокращать ненужные степени свободы нарушением принятых правил и условностей.
Половой диморфизм в антропологии
По утверждению Геодакяна, представления теории пола, об обособленности новой и старой информации на протяжении многих поколений, позволяет объяснить ряд непонятных явлений в антропологии[62]. Так в туркменской популяции методом обобщённого портрета была обнаружена четкая разница по полу — женские портреты укладывались в один тип, а мужские — в два типа[63]. Аналогичное явление наблюдал Р. М. Юсупов в краниологии башкир — женские черепа были близки к угро-финскому типу (в географическом плане это северо-западные соседи современных башкир), а мужские — к алтайскому, казахскому и другим (восточные и юго-восточные соседи)[64]. В удмуртской популяции, дерматоглифика у женщин соответствовала северо-западному типу, а у мужчин — восточно-сибирскому[65]. Л. Г. Кавгазова отметила сходство дерматоглифики болгар с турками, тогда как болгарки были ближе к литовцам.
Женские формы фенотипов показывают исходный этнос, тогда как мужские формы — число источников и направление генных потоков. Факты приведенные выше, показывают угро-финское происхождение удмуртских и башкирских этносов, различающихся по культуре и языку. Четырёхмодальное распределение черепов мужской части популяции, по утверждению В. Геодакяна, объясняется влиянием трёх различных нашествий с юга и востока. Направление генных потоков в этих популяциях — с юго-востока на северо-запад, а для популяции болгар — с юга на север. Он утверждает также, что островная популяция (японская), в полном соответствии с теорией, является мономодальной для обоих полов[62].
Эволюционная теория пола — правила
Экологическое правило дифференциации полов
В оптимальных, стабильных условиях среды, когда нет необходимости в высокой эволюционной пластичности, основные характеристики уменьшаются и имеют минимальное значение, — то есть падает рождаемость (одновременно и смертность) мальчиков, сокращается их разнообразие и разница между мужским и женским полом. Все это снижает эволюционную пластичность популяции. В экстремальных же условиях изменчивой среды, когда для быстрой адаптации требуется высокая эволюционная пластичность, идут обратные процессы: растут, одновременно, рождаемость и смертность (то есть «оборачиваемость») мужского пола, его разнообразие, чётче становится половой диморфизм. Все это повышает эволюционную пластичность популяции.
Правило критерия эволюции признака
Признак эволюционирует — если по нему существует половой диморфизм, и стабилен — когда половой диморфизм отсутствует.
Филогенетическое правило полового диморфизма
«Если по какому-либо признаку существует генотипический популяционный половой диморфизм, то этот признак эволюционирует от женской формы к мужской»[66].
При этом, если дисперсия признака у мужского пола больше, чем у женского — эволюция находится в дивергентной фазе, если дисперсии полов равны — фаза эволюции стационарная, если дисперсия больше у женского пола — то фаза конвергентная. (Филогенетическое правило дисперсии полов)[2].
Правило является частью «Эволюционной теории пола». С точки зрения системного подхода, примененного В. А. Геодакяном в 1965 года к проблеме пола, половой диморфизм рассматривается как следствие асинхронной эволюции полов. Следовательно, половой диморфизм возникает только по эволюционирующим признакам. Это эволюционная «дистанция» между полами, которая появляется с началом эволюции признака и исчезает с её концом. Соответственно, половой диморфизм может быть следствием любого вида отбора, а не только полового, как считал Чарлз Дарвин.
Филогенетическое правило дисперсии полов
Если дисперсия признака у мужского пола больше, чем у женского — эволюция находится в дивергентной фазе, если дисперсии полов равны — фаза эволюции стационарная, если дисперсия больше у женского пола — то фаза конвергентная. Дисперсия — разнообразие признаков у мужского и женского пола.
В раздельнополой популяции, каждый пол имеет свою величину дисперсии — и . Другие параметры — это число особей , соотношение полов и половой диморфизм. Суммарный вклад дисперсии, соотношения полов и полового диморфизма, определяет степень дифференциации полов.



Различают генотипическую и фенотипическую дисперсию полов.
Дисперсия полов в онтогенезе
По аналогии с соотношением полов для разных стадий онтогенеза, можно выделить также первичную, вторичную и третичную дисперсию полов. Поскольку дисперсия связана с признаками, а в зиготе большинство признаков ещё в потенции, под первичной дисперсией надо понимать те потенции, из которых будет реализован признак во взрослой дефинитивной стадии.
- Генотипическая дисперсия
Повышенный уровень мутаций у мужских особей
Поскольку число клеточных делений в сперматогенезе намного больше числа делений в овогенезе и ошибки при репликации и репарации ДНК являются главным источником мутаций для молекулярной эволюции, был сделан вывод, что все это может привести к большей частоте мутаций в половых хромосомах, по сравнению с аутосомами, и было предположено, что самцы служат генератором мутаций, по крайней мере, в эволюции млекопитающих. (Тот факт, что уровень, как спонтанного так и индуцированного мутагенеза у гетерогаметных и у гомогаметных самцов выше чем у самок (для дрозофилы, шелкопряда, млекопитающих, в том числе и человека) был установлен давно и неоднократно[67]). Более высокий уровень точечных мутаций отмечен также у самцов мышей, по сравнению с самками[68].
Наследование родительских признаков
Было обнаружено, что женские потомки наследуют родительские признаки более аддитивно (промежуточное, среднеарифметическое наследование), чем мужские. Различия между самцами и самками мышей наблюдались для относительного веса надпочечников[69], тимуса, половых желез и гипофиза[70], а также генов, ответственных за двигательную активность[71].
Фенотипическая дисперсия
Бо́льшая фенотипическая дисперсия мужского пола является одним из основных положений эволюционной теории пола. Так как фенотипическая дисперсия отражает генотипическую, можно ожидать, что у мужских особей она должна быть шире за счет мутаций и неаддитивного наследования признаков. Степень связи генотипа с фенотипом (норма реакции) также определяет величину фенотипической дисперсии.
Дисперсия полов в филогенезе
Согласно эволюционной теории пола В. А. Геодакяна, дисперсия полов — так же, как и соотношение полов и половой диморфизм, — тесно связана с одной стороны с условиями среды, а с другой — с эволюционной пластичностью, необходимой на данной стадии эволюции признака.
Отрицательная обратная связь регулирующая дисперсию полов популяции
Контроль дисперсии может осуществляться через Х хромосомное наследование и механизм гомо-/гетерозиготности[72]. В генотипе мужского пола все Х хромосомные гены находятся в гемизиготном состоянии и все рецессивные признаки проявятся, тогда как в женском генотипе признаки проявятся только в гомозиготном состоянии. Такие признаки находятся на периферии фенотипического распределения и подвергаются действию отбора (оперативная часть). Гетерозиготные женские генотипы должны находиться в центре распределения (консервативная часть).
В результате скрещивания, мужские генотипы все равно остаются на периферии распределения, не меняя соотношения между оперативной и консервативной частями. Женские же генотипы переходят из гетерозиготного состояния в гомозиготное и наоборот. При этом, гомозиготизация приводит к их переходу с центра на периферию и увеличивает дисперсию, тогда как гетерозиготизация приводит к обратному эффекту. Этот механизм способен регулировать дисперсию и автоматически поддерживает её оптимум. С уменьшением дисперсии, пропорция женских гетерозигот возрастает, что приводит к большей гомозиготизации в следующем поколении. И наоборот, увеличение дисперсии (рост пропорции женских гомозигот) приводит к гетерозиготизации и уменьшению дисперсии (отрицательная обратная связь).
Онтогенетическое правило полового диморфизма
«Если по какому-либо признаку существует популяционный половой диморфизм, то в онтогенезе этот признак меняется, как правило, от женской формы к мужской»[73].
Можно сказать также, что для начальной, ювенильной стадии онтогенеза более характерна женская форма признака, а для дефинитивной, зрелой стадии — мужская. Другими словами, женские формы признаков с возрастом должны, как правило, ослабевать, а мужские формы — усиливаться.
На более тесную связь женского пола с начальной фазой онтогенеза обратил внимание ещё Дарвин. Он писал: «Во всем животном царстве, если мужской и женский пол отличаются друг от друга по внешнему виду, видоизменяется, за редкими исключениями, самец, а не самка, потому что последняя обыкновенно остается схожей с молодыми животными своего вида и с прочими членами всей группы»[74]. Антропологи также отмечали близость женского типа с детским (более грацильные кости, слабовыраженные надбровные дуги, меньшее оволосение тела и др.).
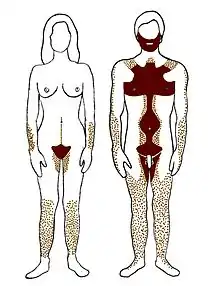
Ярким примером может служить связь степени развития рогов, у разных видов оленей и антилоп, с возрастом их появления у самцов и самок: чем сильнее выражена рогатость у вида в целом, тем в более раннем возрасте появляются рога: сперва у самцов и позже у самок. Онтогенетическое правило полового диморфизма было подтверждено на 16 антропометрических признаках: относительная длина ног, предплечья, 4-го и 2-го пальцев[75], головной индекс, окружность зубной дуги[76][77], эпикантус, горбинка спинки носа, оволосение тела, лица и головы[77], концентрация эритроцитов в крови[75], частота пульса[78], скорость опорожнения желчного пузыря[79], асимметрия мозга, время реакции[80], ощущение горького вкуса фенилтиомочевины и обоняние[75].
Филогенетическое правило реципрокных эффектов
«У реципрокных гибридов по дивергирующим признакам родителей должна доминировать отцовская форма (порода), а по конвергирующим — материнская.»
Тератологическое правило полового диморфизма
«Аномалии развития, имеющие „атавистическую“ природу чаще должны появляться у женского пола, а имеющие „футуристическую“ природу (поиск),— у мужского»[3].
По видовым (и выше рангов общности) признакам (многоклеточность, теплокровность, число органов, план и принципиальное строение тела и др.) половой диморфизм в норме отсутствует. Он наблюдается только в области патологии и выражается в разной частоте появления тех или иных врожденных пороков развития у мужского и женского пола. Классификация врожденных аномалий на «атавистические» (возвраты или остановки развития) и «футуристические» (поиск новых путей) позволяет, в некоторых случаях, проследить в половом диморфизме такие предсказанные эволюционной теорией пола общие тенденции. Например, среди около 2000 новорожденных детей, появившихся на свет с одной почкой, было примерно в два раза больше мальчиков, в то время как среди 4000 детей с тремя почками примерно в 2,5 раза больше девочек. У ланцетников и морских червей (далеких предшественников млекопитающих) в каждом сегменте тела имеется по паре специализированных выделительных органов — метанефридиев. Следовательно, появление трёх почек можно, в определенном смысле, рассматривать как «атавистическую» тенденцию, а одной почки — как «футуристическую». Эта же картина наблюдается среди новорожденных детей со сверхнормативным числом ребер, позвонков, зубов и других органов, претерпевших в процессе эволюции редукцию числа, олигомеризация, — среди них больше девочек. Среди же новорожденных с их нехваткой — наоборот, больше мальчиков.
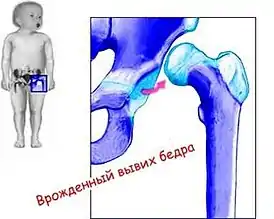
Другая патология — врожденный вывих бедра — встречается у девочек в 4—5 раз чаще, чем у мальчиков[81]. Отметим, что дети с этим дефектом лучше, чем нормальные, бегают на четвереньках и лазят по деревьям. Можно привести также встречающиеся ещё у Дарвина ссылки на сверхнормативные мышцы, которые в 1,5 раза чаще обнаруживаются в трупах мужчин, чем женщин. Или же данные о частоте появления новорожденных с 6-м пальцем, — здесь также число мальчиков в 2 раза превышает число девочек[74].
Правило было также проверено на материале врожденных пороков сердца и магистральных сосудов (32 тыс. случаев)[3]. Было показано, что женские аномалии развития носят характер сохранения эмбриональных особенностей строения сердца, свойственных последним стадиям внутриутробного развития, или же признаков, свойственных видам, стоящим на более низких ступенях эволюционной лестницы (недалекое прошлое) (открытое овальное отверстие в межпредсердной перегородке и Боталлов проток). Элементы «мужских» пороков (стенозы, коарктации, транспозиция магистральных сосудов) имеют «футуристическую» природу (поиск).
Эпидемиологическое правило соотношения полов
Правило устанавливает связь между возрастной и половой эпидемиологией. «Детскими болезнями чаще болеют женщины, болезнями пожилого возраста—мужчины»
Правило соответствия
Если имеется система связанных между собой явлений, в которых можно выделить ориентированные во времени прошлую и будущую формы, то существует соответствие (более тесная связь) между всеми прошлыми формами, с одной стороны, и между будущими — с другой.
Правило соответствия было сформулировано В. А. Геодакяном в 1983 году[82] и проиллюстрировано на примере прошлых и будущих форм признаков в разных явлениях.
В 1866 году был открыт биогенетический закон Геккеля-Мюллера, устанавливающий связь между явлениями филогенеза и онтогенеза (онтогенез — краткое повторение филогенеза).
Если для простоты говорить не об организме в целом, а только об одном его признаке, то явление филогенеза — это динамика (появление и изменение) признака в эволюционном масштабе времени, в истории вида. Явление онтогенеза — это динамика признака в истории жизни особи. Следовательно, закон Геккеля-Мюллера связывает между собой онтогенетическую и филогенетическую динамики признака.
В 1965 году В. А. Геодакяном была открыта закономерность, связывающая явление популяционного полового диморфизма с филогенезом[66]. «Если по какому-либо признаку существует генотипический популяционный половой диморфизм, то этот признак эволюционирует от женской формы к мужской».
В 1983 году им же была теоретически предсказана закономерность, связывающая явление полового диморфизма с онтогенезом[82]. «Если по какому-либо признаку существует популяционный половой диморфизм, то в онтогенезе этот признак меняется, как правило, от женской формы к мужской».
Введем понятия двух форм признака, связанных с вектором времени, в каждом из трёх явлений (филогенеза, онтогенеза и полового диморфизма). В филогенезе признака будем различать его «атавистическую» и «футуристическую» формы, в онтогенезе признака — его «ювенильную» (молодую) и «дефинитивную» (взрослую) формы, а в популяционном половом диморфизме — его «женскую» и «мужскую» формы. Тогда обобщённую закономерность, связывающую явления филогенеза, онтогенеза и полового диморфизма, можно сформулировать как «правило соответствия» между атавистической, ювенильной и женской формами признаков, с одной стороны, и между футуристической, дефинитивной и мужской формами — с другой.
«Правило соответствия» можно распространить и на другие явления, системно связанные с филогенезом и онтогенезом (эволюцией), в которых можно выделить прошлую и будущую формы. Например, явление мутирования (филогенетический процесс возникновения генов), явление доминирования (онтогенетический процесс проявления генов), явление гетерозиса и реципрокных эффектов. На связь между явлениями филогенеза, онтогенеза, мутирования, доминирования и полового диморфизма указывают такие известные факты, как: более высокая степень спонтанных мутаций у мужского пола; более аддитивное наследование родительских признаков потомками женского пола, а значит, более доминантное наследование потомками мужского пола;[83] известные аутосомные гены, проявляющие себя в женском геноме как рецессивные признаки, а в мужском — как доминантные и усиливающиеся в онтогенезе, например ген рогатости—комолости у овец, или ген, вызывающий облысение у человека, а также доминирование отцовской формы над материнской по эволюционирующим (новым) признакам («отцовский эффект»)[84].
Предсказания
Филогенетическое и онтогенетическое правила полового диморфизма, связывающие явление полового диморфизма с динамикой признака в филогенезе и онтогенезе, дают возможность зная одно явление предсказать два других.
Известно, что у далеких филогенетических предшественников человека глаза были расположены латерально, их зрительные поля не перекрывались и каждый глаз был связан только с противоположным полушарием мозга — контралатерально. В процессе эволюции у некоторых позвоночных, в том числе и у предков человека, в связи с приобретением стереоскопического зрения, глаза переместились вперед. Это привело к перекрытию левого и правого зрительных полей и к появлению новых ипсилатеральных связей: левый глаз — левое полушарие, правый глаз — правое. Таким образом, появилась возможность иметь в одном месте зрительную информацию от левого и правого глаза, — для их сопоставления и измерения глубины. Стало быть, ипсилатеральные связи филогенетически более молодые, чем контралатеральные. На основании филогенетического правила, можно предсказать эволюционно более продвинутые ипси-связи у мужского пола, по сравнению с женским, — то есть половой диморфизм по доле ипси/контра волокон в зрительном нерве. На основании онтогенетического правила можно предсказать возрастание доли ипси волокон в онтогенезе. А поскольку зрительно-пространственные способности и объемное воображение тесно связаны со стереоскопией и ипси связями, то становится понятным, почему у мужчин они развиты лучше. Этим объясняются наблюдаемые различия мужчин и женщин в понимании геометрических задач, ориентировании и определении направлений, чтении чертежей и географических карт[85].
Применение тех же правил к обонятельному рецептору человека, приводит к выводу, что в филогенезе обоняние человека, в отличие от зрения, ухудшается. Поскольку, как было показано, у людей с возрастом происходит атрофия обонятельных волокон и их количество в обонятельном нерве неуклонно уменьшается[86][87], то можно предсказать, что их число у женщин должно быть больше, чем у мужчин.
- У большинства видов позвоночных, эволюция которых сопровождалась увеличением размеров, самцы чаще должны быть крупнее самок.
- У многих видов насекомых и паукообразных, которые наоборот мельчали, самцы должны быть мельче самок.
- По всем селекционным признакам, у культурных растений и животных, самцы должны превосходить самок.
- У реципрокных гибридов, по дивергирующим признакам родителей, должна доминировать отцовская форма (порода), а по конвергирующим — материнская.
- Признаки недалекого филогенетического прошлого чаще должны встречаться у особей женского пола, а признаки недалекого возможного будущего — у особей мужского пола.
- Предсказание было подтверждено анализом 31814 больных с врожденными пороками сердца и магистральных сосудов.
- Сверхнормативные мышцы в 1,5 раза чаще обнаруживаются у мужчин, чем у женщин.
- Известно, что относительные размеры мозолистого тела в онтогенезе человека увеличиваются. Это значит, что оно должно быть больше у мужчин и расти в филогенезе.
Критика и отношение к другим теориям
Критика теории пола в целом в литературе отсутствует. Критика отдельных аспектов иногда встречается. Например в книге Л. А. Гаврилова и Н. С. Гавриловой анализируются половые различия в продолжительности жизни[88]. В отношении большей вариабельности признаков у самцов ответственной за их повышенную смертность, авторы отмечают, что «в данной гипотезе не раскрыт конкретный молекулярно-генетический механизм, приводящий к большей продолжительности жизни самок». И там же пишут, что этот недостаток, «может быть в принципе устранён в ходе дальнейшего развития и конкретизации данной гипотезы». Они считают, что предсказание теории о преобладании мужчин среди долгожителей не согласуется с фактами, поскольку, во-первых, «по мере роста продолжительности жизни растут и различия по этому признаку между мужчинами и женщинами» и, во-вторых, «в последние годы в развитых странах наблюдается ускоренное по сравнению с мужчинами снижение смертности женщин старших возрастов»[89]. Они также считают, что «большая продолжительность жизни самок вовсе не является общебиологической закономерностью». Следует отметить, что вывод о большей продолжительности жизни самок у большинства изученных видов был сделан задолго до появления теории пола в целом ряде работ[90][91].
Положения теории о соотношении полов и «Феномен военных лет» обсуждались в работе В. Искрина[92]
Поскольку сам Ч. Дарвин считал, что мужской пол меняется раньше, основное положение концепции В. Геодакяна, что эволюция полов происходит асинхронно, не противоречит теории эволюции Дарвина. В последнее время на Западе даже широко употребляется новый термин англ. «male-driven evolution». Теория В. Геодакяна дополняет и расширяет теорию полового отбора Ч. Дарвина отмечая, что половой диморфизм может возникнуть в результате любого (а не только полового) отбора. А. С. Кондрашов в классификации теорий пола поместил её в категорию «Гипотез немедленного преимущества» (immediate benefit) поскольку отбор среди «дешевых» самцов и мужских гамет более эффективен[93].
Теория В. Геодакяна анализирует процесс половой дифференциации, и поэтому не противоречит многочисленным теориям, пытающимся объяснить возникновение и поддержание полового размножения, поскольку они уделяют основное внимание процессу скрещивания.
Среди теорий раздельнополости, теория пола является более общей, чем например теория Паркера (1972), которая объясняет половую дифференциацию на уровне гамет и только у водных животных[94].
См. также
Примечания
- Жуков Д. А. (2007) Биология поведения. Гуморальные механизмы. СПб.: Речь. С. 369. 466 с.
- Геодакян В. А. (1986) Половой диморфизм. Биол. журн. Армении. 39 № 10, с. 823—834.
- Геодакян В. А., Шерман А. Л. (1970) Экспериментальная хирургия и анестезиология. 32 № 2, с. 18-23.
- Геодакян В. А., Шерман А. Л. (1971) Связь врожденных аномалий развития с полом. Журн. общ. биологии. 32 № 4, с. 417—424.
- Геодакян В. А., Геодакян С. В. (1985) Существует ли отрицательная обратная связь в определении пола? Журнал общей биологии. 46 № 2, с. 201—216.
- Геодакян В. А. (1974) Дифференциальная смертность и норма реакции мужского и женского пола. Журн. общ. биологии. 35 № 3, с. 376—385.
- Геодакян В. А. (1996) Половые хромосомы: для чего они? (Новая концепция). Докл. АН. 346 с. 565—569.
- Геодакян В. А. (1992) Эволюционная логика функциональной асимметрии мозга. Докл. АН. 324 № 6, с. 1327—1331.
- Геодакян В. А., Геодакян К. В. (1997) Новая концепция леворукости. Докл. РАН. 356 № 6, с. 838—842.
- Геодакян В. А. (2005) Эволюционные теории асимметризации организмов, мозга и тела. Успехи физиологических наук. 36 № 1. с. 24-53.
- Trofimova I. Do psychological sex differences reflect evolutionary bi-sexual partitioning? // American Journal of Psychology, 128(4). — 2015. — С. 485—514. — doi:10.5406/amerjpsyc.128.4.0485.
- Публикации
- Хронология и основные события (недоступная ссылка)
- Соколова Е. И. (1992) Два пола зачем и почему? Всероссийский институт повышения квалификации инженерно-педагогических работников и специалистов профтехобразования. С.-Петербург, 41 с.
- Васильченко Г. С. (1977, 2005) Общая сексопатология. Руководство для врачей. М., Медицина.
- Ткаченко А. А., Введенский Г. Е., Дворянчиков Н. В. (2001) Судебная сексология. Руководство для врачей. М., Медицина.
- Ильин Е. П. Дифференциальная психофизиология мужчины и женщины. 2003.
- Жуков Д. А. (2007) Биология поведения. Гуморальные механизмы. С-Пб.: Речь, 2007. 443 с. Архивная копия от 10 сентября 2013 на Wayback Machine
- Палій А. А. Диференціальна психологія. Навчальний посібник. К. Академія Рік видання, 2010, 432 c.
- Кравець В., Говорун Т., Кікінежді О. Ґендерна освіта: теорія та практика: навчальна програма інтегрованого курсу для студентів ІІ-ІV курсів педагогічних спеціальностей. Київ-Тернопіль, 2011, 282 с., с. 104.
- Каган В. Е. Воспитателю о сексологии. М., Педагогика, 1991.
- С. А. Боринская Эволюционная теория пола. Биология в школе. М., Школа-пресс, 1995, № 6, с. 12-18.
- Тюгашев Е. А. (2002) Экономика семьи и домашнего хозяйства. Учебное пособие. Новосибирск: СибУПК, с. 33-38.
- Нартова-Бочавер С. К. (2003) Дифференциальная психология: Учебное пособие. М.: Флинта, Московский психолого-социальный институт. с. 158—179.
- Андреева Т. В. Семейная психология: Учеб. пособие. СПб.: Речь, 2004. 244 с.
- Московский физико-технический институт. Факультет Молекулярной и Биологический Физики (ФМБФ). Курс лекций «Основы биологии» для 1-го курса. Лекция 24 «Эволюционная теория пола. Биотехнологии. Иммунология. Передача сигналов в организме». http://www.fizhim.ru/student/files/biology/biolections/lection24/
- Харьковский национальный университет. Психологический факультет. Кафедра общей психологии. Иванова Е. Ф. Гендерные исследования в психологии. Лекция 11. Исследования гендерных различий организации мозга и когнитивной сферы. Архивированная копия. Дата обращения: 6 июня 2016. Архивировано 3 марта 2016 года.
- Гордон А. (2002) Эволюционная теория пола. Программа «00:30» НТВ, 06.03.2002 http://www.ntv.ru/gordon/archive/907/
- Гордон А. (2002) Эволюционная теория пола-2. Программа «00:30» НТВ, 15.04.2002
- Гордон А. (2003) Теория асимметрии мозга. Программа «00:30» НТВ, 09.12.2003. http://www.gordon.ru/konkurssite/for_print/031209st_p.html Архивная копия от 24 августа 2004 на Wayback Machine
- Katsenelinboĭgen A. Evolutionary Change; Toward a Systemic Theory of Development and Maldevelopment. Newark: Gordon & Breach Publishing Group, 1997.
- Афонькин С. Ю. (2010) Секреты наследственности человека. Корона принт, С.-Пб., 77 стр.
- Белкин А. И. Третий пол. Судьбы пасынков Природы. М.: Олимп, 2000. 432 с.
- Брагина Н. Н., Доброхотова Т. А. (1988) Функциональные асимметрии человека. 2-е изд., М.: Медицина. С. 55. 240 с.
- Баллюзек Ф. В., Сквирский В. Я., Скоробогатов Г. А. (2009) Говорят учёные (на грани сенсаций): о питьевой воде и водопроводе, о лечебном серебре и алкоголе, о гиподинамии и диете, … СПб.
- Высоцкий Д. Л. (2004) Элементы биологических концепций: Теория построения в приложениях и примерах. Новосибирск: Наука, 570 стр., с. 234—242.
- Kondrashov A. S. J. of Heredity, 1993, v. 84, p. 372—387.
- Похиленко А. П. (2012) ОЦІНКА МОРФОЛОГІЧНОЇ МІНЛИВОСТІ ПОПУЛЯЦІЙ MEGAPHYLLUM SJAELANDICUM (Diplopoda, Julida). Екологія та ноосферологія. 23, № 1-2.
- Пантелеев П. А. (2003) О роли гипотез в зоологических исследованиях. Вестник Зоологии, 37, № 2, с. 3-8.
- Раевский П. М., Шерман А. Л. (1976) Значение пола в эпидемиологии злокачественных опухолей (системно-эволюционный подход). В сб.: Математическая обработка медико-бнологической информации. М.: Наука. С. 170—181.
- Разумов В. В. Ещё раз о философии медицины. Фундаментальные исследования, 2011, № 11, с. 433—439. с.438.
- Журавлева М. И., Соболева Т. С. О некоторых гендерных вопросах исследования психологии творчества. МКО, 2005, ч. 1, с. 165—171.
- Кочетова Т. В. Эволюционный подход в социально-психологических исследованиях: половые различия как ведущие детерминанты статусных предпочтений. Социальная психология и общество, 2010, № 1, с. 78-90.
- Trofimova, IN. Are men evolutionary wired to love the “Easy” buttons? Nature Preceedings hdl:10101/npre.2011.5562.1, p. 1–16. (2011).
- Панкратов А. В., Воркунова И. А. Коммуникативные компоненты практического мышления. Образование для XXI века: VIII Международная научная конференция (Москва, 17—19 ноября 2011 г.): Доклады и материалы. Секц. Психологические проблемы образования в XXI веке / Отв. ред. А. Л. Журавлев. — М.: Изд-во Моск. гуманит. ун-та, 2011. — С. 50—54. 92 с.
- Засядь-Волк Ю. В. (2011) Проблема смысла жизни и половой дипсихизм. Идеи и идеалы, 2, № 1(7), с. 9-19.
- Мажуль Л. А. Дивергентная эволюция: от Природы к Культуре (бинарные оппозиции и гендер). «Мир психологии», 2016, № 3 (87), с. 223—238.
- Еремеева В. Д., Хризман Т. П. (1998) Мальчики и девочки — два разных мира. Нейропсихологи — учителям, воспитателям, родителям, школьным психологам. М.: Линка-Пресс. 184 с.
- Геодакян В. А. (1989) Теория дифференциации полов в проблемах человека. Человек в системе наук. М.: Наука. С. 171—189.
- Геодакян В. А. (1991) Эволюционная теория пола. Природа. № 8. С. 60-69.
- Геодакян В. А. (1973) Дифференциальная смертность полов и норма реакции. Биол. журн. Армении. 26 № 6, с. 3—12.
- Vandenberg S. G., McKusick V. A., McKusick A. B. (1962). Twin data in support of the Lyon hypothesis. «Nature». 194 № 4827 505—506.
- Chovanova E., Bergman P., Stukovsky K. (1980) Abstracts of communications of II Congress of European Anthropological Association, Brno, 136.
- Геодакян В. А. (1977) Количество пыльцы как регулятор эволюционной пластичности перекрёстноопыляющихся растений. ДАН СССР, т. 234, № 6. с. 1460—1463.
- Hamilton J. B. (1948). The role of testicular secretions as indicated by the effects of castration in man and by studies of pathological conditions and the short life span associated with maleness. «Recent Progress in Hormone Research» 3 N.Y., Acad. Press, 257—322.
- Stump J. B. (1985). What’s the Difference? How Men and Women Compare. N.Y., William Morrow & Co., Inc.
- Геодакян В. А. (2000) Эволюционные хромосомы и эволюционный половой диморфизм. Известия Академии Наук, Серия Биологическая, № 2, с. 133—148.
- Бондаренко Ю.В., Романов М.М. (1994-12-20). «Особливості статевої мінливості маси тіла у добового молодняка свійських птахів» in Генетика продуктивності тварин (Київ, 20—21 грудня 1994). Тези доповідей Всеукр. ювіл. наук.-практ. конф., присвячен. 90-річчю з дня народж. видатного вченого Колесника М. М.: 36. OCLC 899128304. Дата обращения: 2020-10-19. (укр.)
- Геодакян В. А. (1981) Половой диморфизм и «отцовский эффект». Журн. общ. биологии, 42, № 5, с. 657—668.
- Еремеева В. Д., Хризман Т. П. Мальчики и девочки — два разных мира. Нейропсихологи — учителям, воспитателям, родителям, школьным психологам. М.: Линка-Пресс, 1998. 184 с.
- Trofimova IN. Understanding misunderstanding: a study of sex differences in meaning attribution // Psychological Research. — 2012. — Т. 77(6). — С. 748—760. — doi:10.1007/s00426-012-0462-8.
- Геодакян В. А. (1989) Теория дифференциации полов в проблемах человека. Человек в системе наук. М.: Наука. С. 171—189. 504 с.
- Павловский О. М. (1980) О чем рассказывает обобщённый фотопортрет? Наука и жизнь, № 1, с. 84-90.
- Юсупов Р. М. (1986) О половом диморфизме и значении женских выборок черепов в антропологии. Источники по истории и культуре Башкирии. Уфа, с. 51-56.
- Долинова Н. А. (1989) Дерматоглифика удмуртов. Новые исследования по этногенезу удмуртов. Ижевск, Уро АН СССР. с. 108—122.
- Геодакян В. А. (1965) Роль полов в передаче и преобразовании генетической информации. Пробл. передачи информ. 1 № 1, с. 105—112.
- Kerkis J. (1975) Some Problems of Spontaneous and Induced Mutagenesis in Mammals and Man. Mutation Res. 29 p. 271—280.
- Searle A. G. (1972) Spontaneous frequencies of point mutations in mice. Humangenetik, Bd. 16 № 1/2, p. 33-38.
- Badr F. M., Spickett S. G. (1965) Genetic variation in adrenal weight relative to body weight in mice. Acta Endocrinol. Suppl. 100 p. 92-106.
- Шюлер Л., Бородин П. М., Беляев Д. К. (1976) Проблемы генетики стресса. Генетика. 12 № 12, с. 72-82.
- Бородин П. М., Шюлер Л., Беляев Д. К. (1976) Проблемы генетики стресса. Сообщение I. Генетический анализ поведения мышей в стрессирующей ситуации. Генетика. 12 № 12, с. 62-72.
- Geodakjan V.A. (1987) Feedback Control of Sexual Dimorphism and Dispersion. Towards a New Synthesis in Evolut. Biol. Proc. Intern. Symp. Praha. Czech. Ac. Sci. p. 171—173.
- Геодакян В. А. (1983) Онтогенетическое правило полового диморфизма. ДАН СССР. 269 № 2, с. 477—481.
- Дарвин Ч. (1953) Происхождение человека и половой отбор. Соч. — М.: Изд-во АН СССР. 5 1040 с.
- Харрисон, Дж.; Уайнер, Дж.; Таннер, Дж.; Барникот, Н. Биология человека. — М.: Мир, 1968.
- Гинзбург, В. В. Элементы антропологии для медиков. — Л.: Гос. изд. мед. лит, 1963.
- Рогинский Я. Я., Левин М. Г. Основы антропологии. — М., 1955.
- Пульс // Большая медицинская энциклопедия. — М.: Сов. энциклопедия, 1962. 27 С. 515.
- Желчный пузырь // Большая медицинская энциклопедия, 1978. 8 С. 184.
- Бойко, Е. И. Время реакции человека. — М.: Медицина, 1964.
- Cui W., Ma C., Tang Y., e.a. (2005) Sex Differences in Birth Defects: A Study of Opposite-Sex Twins. Birth Defects Research (Part A) 73 p. 876—880.
- Геодакян В. А. (1983) Онтогенетическое правило полового диморфизма. ДАН СССР 269 № 2, с. 477—481.
- Бородин П. М., Шюлер Л., Беляев Д. К. (1976) Генетика. 12 № 12, с. 62.
- Геодакян В. А. (1981) Половой диморфизм и «отцовский эффект». Журн. общ. биологии, 42 № 5, с. 657—668.
- См., например, «Мужская и женская стратегия ориентирования на местности» // Behavioral Neuroscience.
- Блинков С. М., Глезер И. И. (1964) Мозг человека в цифрах и таблицах. — Л. — 180 с.
- Smith C. G. (1942) Age incidence of atrophy of olfactory nerves in man. J. Comp. Neurol. 77 № 3, p. 589—596.
- Гаврилов Л. А., Гаврилова Н. С. (1991) Биология продолжительности жизни. М.: Наука. 280 с.
- Gavrilov L. A., Nosov V. N. A new trend in human mortality decline: derectangularization of the survival curve. — Age, 1985, 8(3): 93-93.
- Hamilton J. B. The role of testicular secretions as indicated by the effects of castration in man and by studies of pathological conditions and the short life span associated with maleness. Recent Progress in Hormone Research, v. 3, N.Y., Acad. Press, 1948, p. 257—322.
- Комфорт А. (1967) Биология старения. М.: Мир. С. 400.
- Lubachevsky, Boris (2009), Theory of sexes by Geodakian as it is advanced by Iskrin, arΧiv:cs/0607007
- Kondrashov A. S. (1993) J. of Heredity. 84 p. 372—387.
- Parker G. A., Baker, R. R., Smith, V. G. F. (1972) The origin and evolution of gamete dimorphism and the male-female phenomenon. J. Theor. Biol., 36 p. 529.