Тормозный постсинаптический потенциал
Тормозный постсинапти́ческий потенциа́л — это разновидность постсинаптического потенциала, которая приводит к тому, что активность постсинаптического нейрона понижается, и менее вероятным становится возникновение потенциала действия[1].
Противоположностью тормозному постсинаптическому потенциалу является возбуждающий постсинаптический потенциал, который приводит к тому, что активность постсинаптического нейрона повышается, и возникновение потенциала действия становится более вероятным.
Возникновение различных типов постсинаптических потенциалов возможно в различных типах химических синапсов, которые используют секрецию тех или иных нейромедиаторов для обеспечения нейротрансмиссии (межклеточной передачи сигнала).
Тормозящие (ингибирующие) пресинаптические нейроны выделяют в синапс тормозные нейромедиаторы (например, такие, как ГАМК, глицин, серотонин, в зависимости от типа нейрона). Эти тормозные нейромедиаторы затем связываются с соответствующими специфическими «тормозными» постсинаптическими рецепторами. В результате активации этих тормозных рецепторов происходят изменения в активности постсинаптического нейрона, в частности открываются или закрываются ионные каналы (например, каналы ионов хлора в случае ГАМК-А рецептора или каналы ионов калия в случае 5-HT1A-рецептора). Это приводит к изменению электрической проводимости мембраны постсинаптического нейрона. Генерируется электрический ток, который изменяет постсинаптический потенциал — постсинаптическая мембрана становится более электроотрицательной (более отрицательно заряженной). Если исходный потенциал мембраны находится между порогом покоя и порогом возникновения потенциала действия, то в результате воздействия этого ингибирующего потенциала может произойти деполяризация клетки. Тормозные постсинаптические потенциалы также приводят к изменению проницаемости мембраны для ионов хлора, поскольку в результате изменения потенциала мембраны изменяется электростатическая сила, воздействующая на хлорные каналы[2]. Для измерения постсинаптических потенциалов в возбуждающих и тормозных синапсах могут использоваться микроэлектроды.
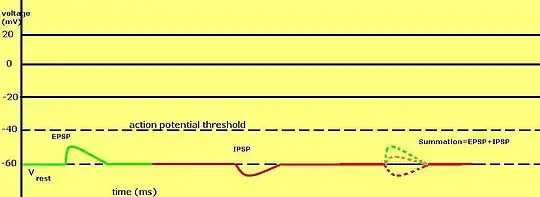
В целом, результирующий постсинаптический потенциал клетки зависит от комбинации факторов: типы и комбинации рецепторов и ионных каналов клетки, одновременно подвергающихся воздействию, характер воздействий (агонистический или антагонистический), исходный постсинаптический потенциал клетки, реверсный потенциал, порог возникновения потенциала действия, проницаемость ионных каналов клетки для тех или иных ионов, а также градиент концентрации ионов внутри и снаружи клетки. Вся эта совокупность факторов в конечном счёте и определяет, будет ли клетка в состоянии возбуждения или в состоянии покоя либо даже угнетения. Тормозные постсинаптические потенциалы всегда направлены на то, чтобы снизить (сделать более электроотрицательным) мембранный потенциал клетки и удержать его ниже порога возникновения потенциала действия. Таким образом, тормозный постсинаптический потенциал может рассматриваться как своего рода «временная гиперполяризация» клетки.[3] Тормозные и возбуждающие постсинаптические потенциалы конкурируют друг с другом на множестве синаптических терминалей нейрона. Их суммация и предопределяет то, будет или не будет потенциал действия, сгенерированный пресинаптической клеткой в конкретном синапсе, повторен (регенерирован) подобным же потенциалом действия на постсинаптической мембране. Эта же суммация всех имеющихся потенциалов предопределяет и то, какой будет реакция постсинаптической клетки на очередной, «ещё один», тормозный или возбуждающий сигнал, не достигающий сам по себе величины потенциала действия. Некоторые типичные нейромедиаторы, вовлечённые в генерацию тормозных постсинаптических потенциалов — это ГАМК и глицин, и — во многих, но не во всех, случаях (в зависимости от типа рецептора) — серотонин.

Компоненты
Типы
Эта система работает таким образом, что[1] тормозные постсинаптические потенциалы суммируются во времени с подпороговыми или надпороговыми возбуждающими потенциалами, в результате чего уменьшается результирующий постсинаптический потенциал. Эквивалентные по модулю возбуждающие (положительные) и тормозные (отрицательные) постсинаптические потенциалы дают в сумме нейтральное состояние, взаимно отменяя действие друг друга на клетку. Баланс между возбуждающими и тормозными постсинаптическими потенциалами очень важен для интеграции клеткой всей поступающей от различных возбуждающих и тормозных синапсов электрической и химической информации.
Дополнительные факторы
Размер нейрона также может влиять на эффект, который оказывает на клетку тормозной постсинаптический потенциал. Простое и мгновенное временное суммирование постсинаптических потенциалов имеет место быть в нейронах сравнительно небольшого размера, тогда как в крупных нейронах большее количество синапсов, метаботропных и ионотропных рецепторов, а также наличие длинных аксонов и большее расстояние от синапсов до тела нейрона позволяет нейронам ещё некоторое время продолжать электрическое и химическое общение с другими нейронами (то есть пребывать в состоянии возбуждения), несмотря на наличие тормозных потенциалов на удалённых от тела синапсах, пока тормозящий сигнал «путешествует» до тела клетки.
Ингибиторные молекулы
ГАМК является весьма распространённым тормозным нейромедиатором (нейромедиатором, воздействие которого приводит к генерации тормозного постсинаптического потенциала) в нервной системе и сетчатке млекопитающих.[1][4] ГАМК-рецепторы являются пентамерами, наиболее часто состоящими из трёх разных субъединиц (α, β, γ), хотя существуют и несколько других субъединиц (δ,ε, θ, π, ρ) и возможных конфигураций ГАМК-рецептора. Открытые каналы селективно проницаемы для ионов хлора или калия (в зависимости от типа рецептора) и позволяют этим ионам проходить через мембрану. Если электрохимический потенциал возникающего при этом ионного тока более отрицателен, чем порог возникновения потенциала действия, то возникающее как следствие этого ионного тока (который сам есть следствие активации ГАМК-рецептора) изменение электрического заряда (потенциала) мембраны и её проводимости приводит к тому, что результирующий постсинаптический потенциал становится ниже (более электроотрицательным), чем порог возникновения потенциала действия, и это снижает вероятность постсинаптического нейрона сгенерировать потенциал действия. Молекулы и рецепторы глицина действуют во многом сходным образом и в нервной системе, и в сетчатке.
Ингибирующие рецепторы
Существует два типа ингибирующих рецепторов:
Ионотропные рецепторы
Ионотропные рецепторы (также известные как открываемые лигандом ионные каналы) играют важную роль в быстрой генерации тормозных постсинаптических потенциалов.[1] Нейромедиатор связывается со специфическим доменом рецептора — так называемым сайтом или доменом связывания лиганда (ligand-binding site or domain) рецептора, расположенным на наружной стороне поверхностной мембраны клетки (обращённым в синаптическую щель). Это приводит к изменению пространственной конфигурации рецептора и к открытию в нём ионного канала, который образуется внутри эндомебранного (проходящего сквозь мембрану) домена рецептора. В результате возникает быстрый входящий или исходящий ток ионов — внутрь или наружу клетки. Ионотропные рецепторы способны продуцировать очень быстрые изменения постсинаптического потенциала — в течение миллисекунд после генерации потенциала пресинаптической клеткой. Ионные каналы способны влиять на амплитуду и временные характеристики потенциала действия клетки в целом. Ионотропные ГАМК-рецепторы, сопряженные с каналами ионов хлора, являются мишенью действия многих лекарств, в частности барбитуратов, бензодиазепинов, ГАМК-аналогов и агонистов, ГАМК-антагонистов, таких, как пикротоксин. Алкоголь также модулирует ионотропные ГАМК-рецепторы.
Метаботропные рецепторы
Метаботропные рецепторы, большинство из которых относятся к семейству G-белок-связанных рецепторов, не содержат встроенных в их структуру ионных каналов. Вместо этого они содержат внеклеточный домен связывания лиганда (ligand-binding domain) и внутриклеточный домен связывания с первичным эффекторным белком, которым наиболее часто бывает G-белок.[1] Связывание агониста с метаботропным рецептором приводит к такому изменению конфигурации рецептора, при котором активируется первичный эффекторный белок. Так, например, в случае G-белка активация связанного с ним рецептора приводит к диссоциации β- и γ- субъединиц G-белка в виде βγ-димера и к активации ими ряда «добавочных» внутриклеточных сигнальных путей (в частности en:GIRK), в то время как активированная α-субъединица G-белка изменяет активность классического аденилатциклазного пути (повышает в случае стимулирующего Gs белка и угнетает в случае ингибирующего Gi). Это, в свою очередь, приводит к изменению внутриклеточной концентрации вторичного посредника — циклического АМФ — повышению, в случае повышения активности аденилатциклазы, или снижению, в случае снижения. А изменение концентрации циклического АМФ влияет на активность цАМФ-зависимой протеинкиназы A — вторичного эффектора. Повышение или снижение активности протеинкиназы А запускает нисходящий эффекторный каскад вплоть до эффекторов N-го порядка. В частности, открываются или закрываются ионные каналы.
Ингибирующие метаботропные рецепторы связаны всегда с ингибирующим подтипом G-белка, то есть с Gi. Таким образом, они угнетают активность аденилатциклазы и снижают концентрацию циклического АМФ, эффективно угнетая тем самым активность протеинкиназы А. Помимо этого, они активируют входящий поток ионов калия через GIRK, активированный βγ-димером G-белка, и угнетают активность кальциевых каналов, что вызывает гиперполяризацию клетки. Именно таким образом устроены метаботропные ГАМК-рецепторы (гетеродимеры R1 и R2 субъединиц). Сходным образом устроен и 5-HT1A-рецептор.
Метаботропные ингибирующие рецепторы генерируют медленные тормозные постсинаптические потенциалы (длительностью от миллисекунд до минут). Они могут активироваться одновременно с ионотропными (с некоторыми типами ионотропных рецепторов они могут формировать «рецепторный дуплет» — гетеродимер) в одном и том же синапсе, что позволяет одному и тому же синапсу генерировать как быстрые, так и медленные тормозные потенциалы.
Значение
Исследования
Примечания
- Purves et al. Neuroscience. 4th ed. Sunderland (MA): Sinauer Associates, Incorporated; 2008.
- Thompson S. M., Gahwiler BH. ACTIVITY-DEPENDENT DISINHIBITION .1. REPETITIVE STIMULATION REDUCES IPSP DRIVING FORCE AND CONDUCTANCE IN THE HIPPOCAMPUS INVITRO (англ.) // Journal of Neurophysiology : journal. — 1989. — Vol. 61. — P. 501—511.
- Levy et al. Principles of Physiology. 4th ed. (PA): Elsevier; 2005.
- Chavas J., Marty A. Coexistence of excitatory and inhibitory GABA synapses in the cerebellar interneuron network (англ.) // Journal of Neuroscience : journal. — 2003. — Vol. 23. — P. 2019—2031.