Рамфоринхи
Рамфори́нхи[1] (лат. Rhamphorhynchus, от др.-греч. ῥάμφος — клюв и ῥύγχος — морда) — род длиннохвостых птерозавров из подсемейства Rhamphorhynchinae семейства Rhamphorhynchidae, живших в юрском периоде (161,2—145,0 млн лет назад) на территории Европы (Португалия, Германия, Великобритания) и Африки (Танзания)[2]. Впервые описан палеонтологом Германом фон Майером в 1847 году. Включает в себя 3 валидных вида. Отличительной чертой рамфоринхов были длинные, укреплённые связками хвосты с характерной ромбовидной кисточкой на конце.
| † Рамфоринхи | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Отряд: Клада: Семейство: † Рамфоринхиды Подсемейство: † Rhamphorhynchinae Род: † Рамфоринхи |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Rhamphorhynchus Meyer, 1847 | |||||||||||||||||||||||||||||||||||||||||
| Виды | |||||||||||||||||||||||||||||||||||||||||
| Геохронология 161,2—145,0 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
Хотя фрагментарные остатки рамфоринхов найдены в Англии, Танзании и Португалии, лучше всего сохранились образцы из Зольнхофенских известняков, Германия. Многие из этих окаменелостей сохранили не только кости, но и отпечатки мягких тканей, таких, как мембрана крыла. Разрозненные зубы, возможно, принадлежавшие рамфоринху, обнаружены в Португалии[3].
Описание
Череп
В своём докладе 1927 года немецкий палеонтолог Фердинанд Бройли утверждал, что обнаружил гребень из тонкой кости высотой в 2 мм, который проходил по большей части длины черепа одного экземпляра, что якобы подтверждалось отпечатками на окружающей породе и небольшими остатками самого гребня[4]. Однако последующие исследования этого образца Петером Велльнхофером в 1975 году и Кристофером Беннеттом в 2002 году с использованием как видимого, так и ультрафиолетового света не нашли никаких следов гребня; учёные пришли к выводу, что Бройли ошибся. Они заключили, что предполагаемый гребень был дефектом сохранения[5][6].
Челюсти рамфоринхов несли острые игольчатые зубы, направленные вперёд, а острый, изогнутый, беззубый клюв указывает, в основном, на рыбную диету; в самом деле, рыба и остатки головоногих часто встречаются среди содержимого желудков рамфоринхов, а также в копролитах[7]. Когда челюсти рамфоринха смыкались, верхний и нижний ряды зубов перекрещивались друг с другом, что также наводит на мысль о рыбной диете[3]. В верхней челюсти птерозавра было 20 зубов, нижняя челюсть несла 14 зубов[3].
Посткраниальный скелет
Скелет рамфоринхов насчитывал 7 шейных, 16 грудных, 2 поясничных, 4—5 крестцовых и 40 и более хвостовых позвонков. Грудина была весьма крупной, четырёхугольной, с хорошо развитым килем и длинным передним выростом. Плечевые кости были относительно короткими, но мощными. Предплечье в полтора раза и более длиннее плеча, пястные кости в полтора-два раза короче плеча и в 18—19 раз короче летательного пальца. Первые (наиболее длинные) фаланrи летательноrо пальца в 2,5 раза длиннее плеча. Предлобные кости тонкие, без расширений, изогнутые под прямым уrлом. Бедренные кости заметно короче плечевых, а большие берцовые равны им или превосходят их по длине. Размеры тела рамфоринхов достигали 40 см и более[8].
История изучения и классификация
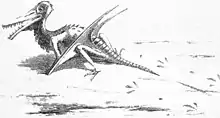
О первом названном образце рамфоринха было доведено до сведения Самуэля Томаса Зёммеринга коллекционером Георгом цу Мюнстером в 1825 году. Зёммеринг пришёл к выводу, что образец принадлежал древней птице. Когда дальнейшее препарирование вскрыло у образца зубы, Мюнстер отправил слепок профессору Георгу Августу Гольдфусу, который определил животное как птерозавра. Как и большинство птерозавров, описанных в середине XIX века, рамфоринхи изначально считались разновидностью птеродактиля. Тем не менее, многие учёные того времени ошибочно называли птеродактиля Ornithocephalus[9]. Поэтому данный образец первоначально был назван Ornithocephalus Münsteri. Об этом впервые упомянул сам Мюнстер в 1830 году[10]. Тем не менее, описание, которое сделало название валидным, было дано в статье Гольдфуса 1831 года, дополнявшей короткую статью Мюнстера[11]. Впоследствии МКЗН запретила использование нестандартных символов латиницы, таких, как ü, в написании научных названий, и написание münsteri было исправлено на muensteri Ричардом Лидеккером в 1888 году.
В 1839 году Мюнстер описал другой вид, который он считал принадлежавшим к Ornithocephalus (птеродактилю), с характерным длинным хвостом. Он назвал его Ornithocephalus longicaudus, что означает «длинный хвост», чтобы отличать его от образцов с короткими хвостами (настоящих птеродактилей)[12].
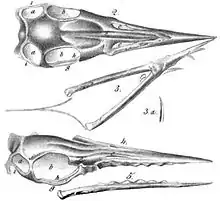
В 1845 году Герман фон Майер официально исправил название вида Ornithocephalus münsteri на Pterodactylus münsteri, поскольку название птеродактиль на тот момент было уже признано и имело приоритет над Ornithocephalus[13]. В своей следующей работе 1846 году с описанием нового вида длиннохвостого «Pterodactyl», фон Майер пришёл к выводу, что длиннохвостые формы достаточно отличаются от короткохвостых форм для того, чтобы разместить их в подрод, и назвал новый вид Pterodactylus (Rhamphorhynchus) gemmingi в честь владельца образца коллекционера Карла Эминга фон Гемминга, который позже продал образец Музею Тейлора в Харлеме[14]. В 1847 году фон Майер «повысил» таксономический статус рамфоринха до полноценного рода и официально включил в него два длиннохвостых вида, известных на то время — Р. longicaudus и Р. gemmingi[15]. Типовым видом рамфоринха стал R. longicaudus, его типовой экземпляр также был продан Музею Тейлора, где размещается до сих пор под индексом TM 6924.
Оригинальный вид, Pterodactylus muensteri, оставался неверно классифицированным до 1861 года, когда Ричард Оуэн опубликовал книгу с переоценкой, где дал виду название Rhamphorhynchus münsteri[16].
Типовой образец R. muensteri, описанный Мюнстером и Гольдфусом, был утерян во время Второй мировой войны. Если есть возможность, в случае утраты типового образца или слишком плохого его состояния, назначается новый типовой образец, или неотип. В своём обзоре 1975 года Петер Велльнхофер отказался назначить неотип, поскольку ряд высококачественных слепков оригинального образца был всё ещё доступен среди музейных коллекций[17]. Они служат пластотипами.
К 1990-м годам, после проведённого Велльнхофером уплотнения многих ранее названных видов, были признаны 5 видов из Зольнхофенских известняков Германии и несколько других из Португалии, Африки и Великобритании на основе фрагментарных остатков[5][17]. Большинство зольнхофенских образцов немного отличались друг от друга размерами, а также признаками, зависящими от размера, например, длиной черепа[17].
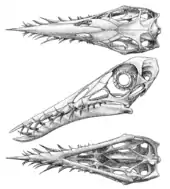
В 1995 году исследователь птерозавров Кристофер Беннетт опубликовал обширный обзор признанных в настоящее время немецких видов. Он пришёл к выводу, что все якобы отдельные зольнхофенские виды на самом деле — возрастные группы одного вида R. muensteri, представляющие особей разного возраста, молодняка в меньшем количестве, взрослых особей — в бо́льшем. Обзор Беннетта не учитывал образцы из Великобритании и Африки, но он предположил, что их следует рассматривать в качестве неопределённых членов семейства Rhamphorhynchidae, а не обязательно как виды самого рамфоринха. Несмотря на сокращение рода к одному виду, типовым видом оставался R. longicaudus[17].
Приведённая ниже кладограмма отражает результат филогенетического анализа, проведённого Андресом и Майерсом в 2013 году[18].
| Breviquartossa |
| |||||||||||||||||||||||||||||||||||||||||||||
Палеобиология


Традиционно, большой разброс в размерах между образцами рамфоринхов объяснялся видовой дифференциацией. Однако, в статье 1995 года Беннетт утверждал, что эти «виды» на самом деле представляют собой возрастные классы одного вида, Rhamphorhynchus muensteri, от птенцов до взрослых. Исходя из этой интерпретации, Беннетт обнаружил несколько существенных изменений, которые происходили у R. muensteri с возрастом[17].
Подростковые особи рамфоринхов имели относительно короткие черепа с большими глазницами. Беззубые кончики клювов у подростков были короче, чем у взрослых; закруглённые, тупые кончики нижних челюстей становились длинными и тонкими по мере роста животных. Взрослые рамфоринхи несли мощный, загнутый вверх «крюк» на нижней челюсти. Количество зубов оставалось постоянным как у подростков, так и у взрослых особей, хотя зубы становились более короткими и толстыми, возможно, приспосабливаясь к более крупной и крепкой добыче. Тазовый и плечевой пояса срастались по мере роста животных; полное срастание грудины происходило к концу первого года жизни[17].
Форма хвостового балансира также меняется в различных возрастных классах. У молодого поколения балансир был маленьким и почти овальным, или, точнее, ланцетовидным. По мере взросления, форма балансира становилась ромбовидной, а у самых крупных особей — треугольной[17].

Самый маленький известный образец рамфоринха имел размах крыла всего 290 мм; тем не менее, вполне вероятно, что даже такая маленькая особь могла летать. Беннетт исследовал две возможности: все рамфоринхи были птенцовыми птерозаврами, что требовало некоторого периода родительской заботы, или выводковыми, чьи птенцы появлялись на свет, уже будучи способными летать. Беннетт предположил, что во втором случае их кладки будут небольшими, с одним или двумя отложенными яйцами, чтобы компенсировать относительно большой размер птенцов. Учёный не стал развивать тему, какими именно были птерозавры, хотя открытие эмбриона птерозавра (Avgodectes) с сильно окостенелыми костями позволило предположить, что в целом птерозавры были выводковыми, способными летать сразу после вылупления и не требующими большой родительской заботы[19]. Эта теория была оспорена гистологическим исследованием рамфоринхов, которое показало, что первоначальный быстрый рост сменялся длительным периодом медленного роста[20].
Метаболизм

Определив, что образцы рамфоринхов вписываются в дискретные возрастные классы, Беннетт смог оценить темпы роста животных в течение одного года, сравнивая размеры однолетних особей с двухлетними. Он обнаружил, что средние темпы роста в течение первого года жизни для рамфоринхов были от 130 % до 173 %, что несколько быстрее, чем темпы роста у аллигаторов. Вероятно, рост значительно замедлялся после половой зрелости, так что достижение максимального размера занимало не более трёх лет[17].
Подобные темпы роста значительно уступают тем, которые наблюдались у крупных представителей подотряда птеродактилей, таких, как птеранодоны, которые достигали своего максимального размера через один год жизни. Кроме того, птеродактили имели детерминированный рост, что означает, что животные достигли фиксированного максимального взрослого размера и переставали расти. Предыдущие предположения о быстром темпе роста у рамфоринхов были основаны на предположении, что они должны быть теплокровными и поддерживающими активный полёт. Теплокровные животные, такие, как современные птицы и летучие мыши, как правило, показывают быстрый рост до определённого размера и последующее прекращение роста. Поскольку никаких доказательств этому у рамфоринхов нет, Беннетт считал, что его доводы указывают на эктотермический метаболизм, хотя он рекомендовал продолжать исследования в этом направлении. Холоднокровные рамфоринхи, по предположению Беннетта, грелись на солнце или разогревали мышцы, чтобы накопить достаточно энергии для короткого полёта, и охлаждались до температуры окружающей среды, будучи неактивными, чтобы сохранить энергию, подобно современным рептилиям[17].
Плавание
Хотя рамфоринхи часто изображаются как воздушные ихтиофаги, последние данные свидетельствуют о том, что они охотились подобно современным морским птицам, ныряя за жертвой в воду. Как некоторые птеранодонтиды, они отличались наличием дельтопекторального гребня в форме топорика, коротким торсом и короткими ногами — всеми признаками у птерозавров, связанными с взлётом с поверхности воды. Ступни рамфоринхов были широкими и большими, полезными для сильного толчка, и указывали на водный образ жизни этих птерозавров[21].
Половой диморфизм
Петер Велльнхофер (1975) признавал две различные группы среди взрослых представителей Rhamphorhynchus muensteri, отличающиеся пропорциями шеи, крыла и задних конечностей, но особенно в соотношении длины черепа к длине плечевой кости. Исследователь отмечал, что эти две группы образцов обнаруживают примерное соотношение 1:1, и интерпретировал их как разнополые[5]. Беннетт протестировал рамфоринха на половой диморфизм, проведя статистический анализ, и обнаружил, что образцы действительно образовывали группы среди множеств малого и большого поголовья. Тем не менее, без какого-либо известного изменения формы костей или мягких тканей (морфологических различий), он не посчитал факт полового диморфизма доказанным[17].
Ориентация головы
В 2003 году группа исследователей во главе с Лоренсом Уитмером изучала анатомию мозга нескольких видов птерозавров, в том числе Rhamphorhynchus muensteri, используя эндокраны мозга, полученные с помощью томографии ископаемых черепов. Используя сравнение с современными животными, учёные смогли оценить различные физические признаки птерозавров, в том числе примерную ориентацию головы во время полёта и координацию мышц крыла. Уитмер и его команда обнаружили, что рамфоринхи держали голову параллельно земле из-за расположения костного лабиринта внутреннего уха, который помогает удерживать баланс. Представители отряда птеродактилей, наоборот, обычно держали головы под уклоном, как в полёте, так и на земле[22].
Модели повседневной активности
Сравнение склеротикальных колец рамфоринхов и современных птиц и рептилий указывает на вероятный ночной образ жизни этих птерозавров, которые, возможно, показывали модели поведения, аналогичные современным ночным морским птицам. Это также может означать, что рамфоринхи делили экологические ниши с дневными птеродактилоидами, такими, как скафогнаты и птеродактили[23].

Экология
Было обнаружено несколько известняковых плит, в которых окаменелые рамфоринхи находятся в тесной связи с ганоидными рыбами Aspidorhynchus. В одном из этих образцов челюсти Aspidorhynchus проходят через крыло рамфоринха; остатки мелкой рыбы, возможно, Leptolepides, находятся в горле рамфоринха. Эта плита иллюстрирует два уровня хищничества — один для рамфоринха и один для Aspidorhynchus. В описании 2012 года авторы предположили, что рамфоринх поймал Leptolepides во время плавания. В то время, пока рыба продвигалась по пищеводу птерозавра, он сам был атакован большим Aspidorhynchus, который случайно проткнул мембрану крыла рамфоринха своим острым носом. Его зубы застряли в пикнофибрах мембраны, и пока рыба билась в попытках освободиться, левое крыло рамфоринха отогнулось назад в искажённом положении, которое запечатлела окаменелость. Стычка привела к гибели обоих животных, скорее всего, потому, что они утонули и попали в бескислородную среду. Они смогли сохраниться вместе, поскольку рыба придавила своим весом гораздо более лёгкого рамфоринха[24].
Виды
По данным сайта Fossilworks, на декабрь 2016 года в род включают 3 вымерших вида[25]:
- Rhamphorhynchus etchesi O'Sullivan & Martill, 2015
- Rhamphorhynchus jessoni Lydekker, 1890
- Rhamphorhynchus muensteri (Goldfuss, 1831)
Ещё один биномен входит в состав рода в статусе nomen dubium — Rhamphorhynchus tendagurensis Reck, 1931[25].
Синонимы
Было названо большое число видов рамфоринхов, которые в настоящее время большинством исследователей принимаются за возрастные классы Rhamphorhynchus muensteri[26]:
|
|
Галерея
 Образец рамфоринха, Хьюстонский музей естественных наук
Образец рамфоринха, Хьюстонский музей естественных наук Голотип R. gemmingi, Музей Тейлора, Харлем
Голотип R. gemmingi, Музей Тейлора, Харлем Образец, формально причисленный к R. intermedius с хвостовым балансиром
Образец, формально причисленный к R. intermedius с хвостовым балансиром Слепок безголового образца R. muensteri, изначально соотнесённого с R. longicaudus, Оксфордский музей естественной истории. Голова была реконструирована для слепка
Слепок безголового образца R. muensteri, изначально соотнесённого с R. longicaudus, Оксфордский музей естественной истории. Голова была реконструирована для слепка
См. также
Примечания
- Татаринов Л. П. Очерки по эволюции рептилий. Архозавры и зверообразные. — М. : ГЕОС, 2009. — С. 177. — 377 с. : ил. — (Труды ПИН РАН ; т. 291). — 600 экз. — ISBN 978-5-89118-461-9.
- Rhamphorhynchus (англ.). Paleobiology Database. (Дата обращения: 19 мая 2016)
- Rhamphorhynchus // The Illustrated Directory of Dinosaurs and Other Prehistoric Creatures / Cranfield, Ingrid (ed.). — London: Salamander Books, Ltd, 2000. — P. 302—305.
- Broili, F. Ein Exemplar von Rhamphorhynchus mit Resten von Schwimmhaut (нем.) // Sitzungs-Berichte der bayerischen Akademie der Wissenschaften mathematisch naturwissenschaftlichen Abteilung. — 1927. — S. 29—48.
- Wellnhofer P. Die Rhamphorhynchoidea (Pterosauria) der Oberjura-Plattenkalke Süddeutschlands (нем.) // Palaeontographica A. — 1975. — Bd. 148, 149. — S. 1—33, 132—186; 1—30.
- Bennett, S.C. Soft tissue preservation of the cranial crest of the pterosaur Germanodactylus from Solnhofen (англ.) // Journal of Vertebrate Paleontology. — 2002. — Vol. 22, no. 1. — P. 43—48. — doi:10.1671/0272-4634(2002)022[0043:STPOTC]2.0.CO;2.
- Frey, E., Tischlinger, H. The Late Jurassic pterosaur Rhamphorhynchus, a frequent victim of the ganoid fish Aspidorhynchus? (англ.) // PLOS ONE. — 2012. — Vol. 7, no. 3. — P. e31945. — doi:10.1371/journal.pone.0031945. — PMID 22412850.
- Основы палеонтологии (в 15 томах) / под ред. Ю. А. Орлова. — М.: Издательство Академии наук СССР, 1959—1963. — Т. 12. Земноводные, пресмыкающиеся и птицы. — С. 597.
- von Sömmerring, S. T. Über einen Ornithocephalus oder über das unbekannten Thier der Vorwelt, dessen Fossiles Gerippe Collini im 5. Bande der Actorum Academiae Theodoro-Palatinae nebst einer Abbildung in natürlicher Grösse im Jahre 1784 beschrieb, und welches Gerippe sich gegenwärtig in der Naturalien-Sammlung der königlichen Akademie der Wissenschaften zu München befindet (нем.) // Denkschriften der königlichen bayerischen Akademie der Wissenschaften. — München: mathematisch-physikalische Classe, 1812. — Bd. 3. — S. 89—158.
- Münster, G. Graf zu. Nachtrag zu der Abhandlung des Professor Goldfuss über den Ornithocephalus Münsteri (Goldf.) (нем.). — Bayreuth, 1830. — S. 8.
- Goldfuss G. A. Beiträge zur Kenntnis verschiedener Reptilien der Vorwelt (нем.) // Nova Acta Academiae Caesareae Leopoldino-Carolinae Germanicae Naturae Curiosorum. — 1831. — Bd. 15. — S. 61—128.
- Münster, G. G. Ueber einige neue Versteinerungen in der lithographischen Schiefer von Baiern (нем.) // Neues Jahrbuch für Mineralogie, Geologie, und Palaeontologie. — 1839. — S. 676—672.
- Meyer, H. von. System der fossilen Saurier [Taxonomy of fossil saurians] (нем.) // Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde. — 1845. — S. 278—285.
- Meyer, H. von. Pterodactylus (Rhamphorhynchus) gemmingi aus dem Kalkschiefer von Solenhofen (нем.) // Palaeontographica. — 1846. — Bd. 1. — S. 1—20.
- Meyer, H. von. Homeosaurus maximiliani und Rhamphorhynchus (Pterodactylus) longicaudus, zwei fossile Reptilien aus der Kalkschiefer von Solenhofen (нем.). — Frankfurt. — Bd. 4X. — S. 22.
- Owen R. Palaeontology, or a Systematic Summary of Extinct Animals and their Geological Relations. — Edinburgh: Adam and Charles Black, 1861. — 463 p.
- Bennett S. C. A statistical study of Rhamphorhynchus from the Solnhofen Limestone of Germany: Year-classes of a single large species (англ.) // Journal of Vertebrate Paleontology. — Vol. 69. — P. 569—580.
- Andres B., Myers T. S. Lone Star Pterosaurs (англ.) // Earth and Environmental Science Transactions of the Royal Society of Edinburgh. — 2013. — Vol. 103. — P. 3—4. — doi:10.1017/S1755691013000303.
- Wang, X., Zhou, Z. Pterosaur embryo from the Early Cretaceous (англ.) // Nature. — 2004. — Vol. 429 (6992), no. 623. — doi:10.1038/429621a.
- Prondvai, E.; Stein, K.; Ősi, A.; Sander, M. P.; Soares, Daphne, ed. Life history of Rhamphorhynchus inferred from bone histology and the diversity of pterosaurian growth strategies (англ.) // PLoS ONE. — 2012. — Vol. 7, no. 2. — P. e31392. — doi:10.1371/journal.pone.0031392.
- Witton, M. P. Were early pterosaurs inept terrestrial locomotors? (англ.) // PeerJ. — 2015. — Vol. 3. — P. e1018. — doi:10.7717/peerj.1018.
- Witmer, L.M., S. Chatterjee, J. Franzosa, T. Rowe, R. C. Ridgely. Neuroanatomy and vestibular apparatus of pterosaurs: Implications for flight, posture, and behavior (англ.) // Integrative and Comparative Biology : Annual Meeting of the Society of Integrative and Comparative Biology, New Orleans, LA. — 2004. — Vol. 43, no. 6. — P. 832.
- Schmitz, L.; Motani, R. Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology (англ.) // Science. — 2011. — Vol. 332, no. 6030. — P. 705—708. — doi:10.1126/science.1200043.
- Frey, E., Tischlinger, H. The Late Jurassic pterosaur Rhamphorhynchus, a frequent victim of the ganoid fish Aspidorhynchus? (англ.) // PLoS ONE. — 2012. — Vol. 7, no. 3. — P. e31945. — doi:10.1371/journal.pone.0031945.
- †Rhamphorhynchus (англ.) информация на сайте Fossilworks. (Дата обращения: 1 декабря 2016).
- †Rhamphorhynchus muensteri (англ.) информация на сайте Fossilworks. (Дата обращения: 1 декабря 2016).
| Таксономия |
|---|