Криптомицеты
Криптомице́ты[3] (лат. Cryptomycota), или розелли́ды[4] (лат. Rozellida), — тип[5] родственных грибам протистов из группы опистоконт. Все представители — облигатные внутриклеточные паразиты или паразитоиды эукариотических организмов. По состоянию на 2015 год таксон включает 3 описанных рода: ''Rozella'' (26 видов), Paramicrosporidium (2 вида) и Nucleophaga (1 вид). Кроме того к криптомицетам относят большое количество последовательностей генов рибосомальных РНК, полученных с помощью методов метагеномики[1].
| Криптомицеты | ||||||||
|---|---|---|---|---|---|---|---|---|
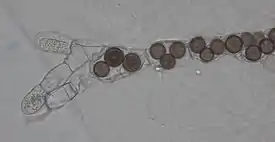 Rozella allomycis в клетках хитридиомицета Allomyces | ||||||||
| Научная классификация | ||||||||
|
Домен: Клада: Obazoa Клада: Клада: Nucletmycea Клада: Opisthosporidia Тип: Криптомицеты |
||||||||
| Международное научное название | ||||||||
| Cryptomycota Jones, 2011 | ||||||||
| Синонимы | ||||||||
| ||||||||
Биология
Все описанные виды роззелид являются внутриклеточными паразитами или паразитоидами. Представители рода Rozella поражают водные формы грибов и оомицетов. Вид R. coleochaetis был обнаружен в зеленой водоросли ''Coleochaete''[6]. Paramicrosporidium и Nucleophaga инфицируют представителей амёбозой, развиваясь внутри ядер своих хозяев[2][7].
Среди клеток криптомицет, выделенных из проб окружающий среды с помощью методики TSA-FISH и не принадлежащих к описанным видам, были обнаружены формы, образующие ассоциации с другими организмами, в первую очередь с диатомеями. Авторы данного исследования предположили, что эти криптомицеты могут вести свободноживущий эпибиотический образ жизни[8]. Однако не исключено, что данные клетки являются инвазивными цистами, прикрепленными к клеткам организма-хозяина (такая стадия жизненного цикла характерна для представителей Rozella)[2].
Жизненный цикл
У розеллид описаны два принципиально различных варианта жизненного цикла. Первый характерен для представителей рода Rozella: подвижные зооспоры контактируют с клеткой хозяина и инцистируются. Циста формирует трубку, прорастающую в клетку хозяина, а также крупную вакуоль на заднем конце. За счет расширения последней происходит внедрение амёбоидного протопласта паразита из цисты в заражаемую клетку. Паразит активно растет и питается, в том числе с помощью фагоцитоза. Затем в клетке паразита начинается деление ядер, и образуется спорангий, в котором формируется большое число новых зооспор, выходящих во внешнюю среду для заражения других клеток или других особей. Кроме того, некоторые виды Rozella могу формировать покоящиеся споры, предназначенные для переживания неблагоприятных условий[6].
Другой вариант жизненного цикла характерен представителям родов Paramicrosporidium и Nucleophaga. Неподвижные споры паразитов фагоцитируются амёбами и инвазируют ядра последних. Паразиты активно растут и питаются, после чего превращаются в одноклеточные спорангии, в которых формируются новые споры. На протяжении всего развития паразитов ядерная оболочка амёб остается интактной. После того как весь объем ядра оказывается занят паразитами, происходит разрушение клетки-хозяина и выход спор во внешнюю среду[2][7].
Особенности строения клетки
Размер клеток розеллид колеблется от 1 до 10 микрометров, в зависимости от стадии жизненного цикла и видовой принадлежности. Все стадии жизненного цикла Rozella, за исключением зрелых покоящихся спор, лишены хитиновой клеточной стенки и обладают стандартным набором клеточных органелл. Зооспоры Rozella несут одиночный жгутик, расположенный на заднем конце клетки[6]. Споры Paramicrosporidium и Nucleophaga во многом похожи на споры микроспоридий, имеют овальную или округлую форму, покрыты хитиновой и белковой оболочками, лишены жгутика и митохондрий. У Paramicrosporidium обнаружены структуры, напоминающие компоненты аппарата экструзии микроспоридий: полярный филомент и якорный диск[2].
Метаболизм
По состоянию на 2015 год данные о метаболизме доступны только для вида Rozella allomycis, геном которого был расшифрован. Митохондриальный геном данного вида демонстрирует значительную степень редукции и представлен кольцевой молекулой размером около 12 тысяч пар нуклеотидов, кодирующей шесть белков — компонентов дыхательной цепи переноса электронов. При этом, у R. allomycis отсутствуют комплекс I данной системы. В геноме у этого вида также обнаружены все компоненты цикла Кребса, что свидетельствует о возможности данного организма продуцировать АТФ с помощью окислительного фосфорилирования. При этом у R. allomycis были обнаружны уникальные элементы метаболической системы, характерные для микроспоридий: альтернативная дыхательная цепь, представленная цитоплазматической и митохондриальной формами НАДН-дегидрогеназы и альтернативной оксидазой, а также нуклеотидные переносчики, используемые микроспоридиями для прямого транспорта АТФ из цитоплазмы клетки-хозяина[9].
Систематическое положение: история и современные представления
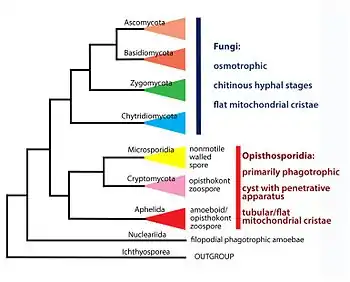
Род Rozella был описан в 1872 году французским ботаником Максимом Корню[11]. Данные организмы считались хитридиевыми грибами до публикации результатов молекулярно-филогенетических исследований Lara c соавторами 2010 года. Авторы продемонстрировали, что Rozella вместе с кладой LKM11, представленной последовательностями генов, выделенных с помощью методов метагеномики, формирует отдельный монофилетический таксон. Авторы определили данную кладу, как наиболее базальную ветвь в составе грибов и дали ей название Rozellida[1].
В 2011 году Jones с соавторами опубликовали формальное таксономическое описание данной группы, где присвоили ей ранг типа и название Cryptomycota[5]. В 2013 году James с соавторами были опубликованы результаты расшифровки генома Rozella allomycis, где было показано, что криптомицеты разделяют множество уникальных черт с микроспоридиями (родственной грибам группой внутриклеточных паразитов), и именно микроспоридии являются ближайшими родственниками розеллид[9].
В 2014 году Corsaro с соавторами описали новый вид криптомицет: Paramicrosporidium, принципиально отличающийся морфологически от Rozella и во многом схожий с микроспоридиями. На основании этого авторы дали новую таксономическую характеристику криптомицет, а также предложили новое название группы: Rozellomycota[2]. В этом же году Karpov с соавторами продемонстрировали, что криптомицеты вместе с микроспоридиями и афелидами (таксон родственных грибам паразитоидов водорослей) являются сестринскими группами и все три клады, в свою очередь, формируют сестринский таксон по отношению к грибам[10][12].
Примечания
- Lara E., Moreira D., López-García P. The environmental clade LKM11 and Rozella form the deepest branching clade of Fungi // Protist. — 2010. — Vol. 161. — P. 116—121. — doi:10.1016/j.protis.2009.06.005.
- Corsaro D., Walochnik J., Venditti D., Steinmann J., Müller K-H., Michel R. Microsporidia-like parasites of amoebae belong to the early fungal lineage Rozellomycota // Parasitol Res. — 2014. — Vol. 113. — P. 1909—1918. — doi:10.1007/s00436-014-3838-4.
- Алёшин В.В. Филогения беспозвоночных в свете молекулярных данных: перспективы завершения филогенетики как науки // Труды зоологического института РАН. — 2013. — № приложение 2. — С. 9—38.
- Леонтьев Д. В. Общая биология: Система органического мира. — Харьков: Харьковская государственная зооветеринарная академия, 2014. — С. 66.
- Jones M. D., Richards T. A., Hawksworth D. L., Bass D. Validation and justification of the phylum name Cryptomycota phyl. nov. // IMA Fungus.. — 2011. — Vol. 2. — P. 173—175. — doi:10.5598/imafungus.2011.02.02.08.
- Gleason, F. H., Carney, L. T., Lilje, O., and Glockling, S. L. Ecological potentials of species of Rozella (Cryptomycota) // Fungal Ecol.. — 2012. — Vol. 5. — P. 651–656. — doi:10.1016/j.funeco.2012.05.003.
- Corsaro D, Walochnik J, Venditti D, Müller KD, Hauröder B, Michel R. Rediscovery of Nucleophaga amoebae, a novel member of the Rozellomycota // Parasitol Res.. — 2014. — Vol. 113. — P. 4491–4498. — doi:10.1007/s00436-014-4138-8.
- Jones MDM, Forn I, Gadelha C, Egan MJ, Bass D, Massana R, Richards TA. Discovery of novel intermediate forms redefines the fungal tree of life (англ.) // Nature. — 2011. — Vol. 474. — P. 200–203. — doi:10.1038/nature09984.
- James, T. Y., Pelin, A., Bonen, L., Ahrendt, S., Sain, D., Corradi, N., et al. Shared signatures of parasitism and phylogenomics unite Cryptomycota and Microsporidia. (англ.) // Current Biology. — Cell Press, 2013. — Vol. 23. — P. 1548-1553. — doi:10.1016/j.cub.2013.06.057.
- Karpov Sergey, Mamkaeva Maria A, Aleoshin Vladimir, Nassonova Elena, Lilje Osu, Gleason Frank H. Morphology, phylogeny and ecology of the aphelids (Aphelidea, Opisthokonta) and proposal for the new superphylum Opisthosporidia // Frontiers in Microbiology. — 2014. — Vol. 5, № 00112. — doi:10.3389/fmicb.2014.00112.
- Cornu M. Monographie des Saprolegniées (фр.) // Annales des Sciences Naturelles, Botanique. — 1872. — Т. 15, № 5. — С. 1—198 (see p. 148).
- Karpov SA, Mamkaeva MA, Benzerara K, Moreira D, López-García P. Molecular phylogeny and ultrastructure of Aphelidium aff. melosirae (Aphelida, Opisthosporidia) // Protist.. — 2014. — Vol. 4, № 165. — P. 512-526. — doi:10.1016/j.protis.2014.05.003.