Альфа-субъединица G-белка
Гуанин-нуклеотид-связывающие белки (так называемые G-белки) — мембранно-связанные гетеротримерные белки, состоящие из трёх субъединиц, называемых «альфа», «бета» и «гамма» (Gα, Gβ, Gγ).[1] Гетеротримерные G-белки и связанные с ними G-белок-связанные рецепторы (GPCR) являются одной из самых распространённых внутриклеточных сигнальных систем в клетках животных, особенно млекопитающих. Они регулируют очень широкий круг процессов, таких, как сенсорные ощущения (зрение, вкус, обоняние), рост, развитие, дифференцировку и размножение клеток, секрецию гормонов[2].
| Альфа-субъединица G-белка | |
|---|---|
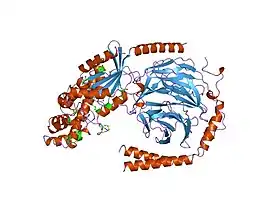 гетеротримерный химерный комплекс Gtα/Giα и субъединица Gβγ | |
| Идентификаторы | |
| Символ | G-alpha |
| Pfam | PF00503 |
| Pfam clan | CL0023 |
| InterPro | IPR001019 |
| SCOP | 1gia |
| SUPERFAMILY | 1gia |
| CDD | cd00066 |
| Доступные структуры белков | |
| Pfam | структуры |
| PDB | RCSB PDB; PDBe; PDBj |
| PDBsum | 3D-модель |
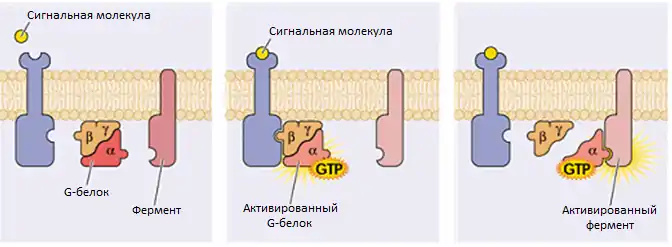
На клеточной поверхности связывание гормонов и нейротрансмиттеров с GPCR активирует рецептор, вызывая конформационные переходы, которые в свою очередь активируют G-белок, расположенный на внутренней поверхности клеточной мембраны. Активация G-белка заставляет ГДФ отсоединиться от его α-субъединицы, а её место занимает ГТФ. Связывание ГТФ вновь приводит к изменению конформации α-субъединицы, что позволяет связанному с рецептором гетеротримеру G-белка (неактивная форма) отсоединиться от него и диссоциировать на активную ГТФ-связанную α-субъединицу и βγ-гетеродимер. α-субъединица и βγ-гетеродимер активируют расположенные ниже эффекторы, таки как аденилатциклаза, фосфодиэстераза, фосфолипазу C и ионные каналы. Эти эффекторы в свою очередь повышают или понижают внутриклеточные концентрации вторичных мессенджеров, например: цАМФ, диацилглицерол, натрий или кальций, которые в конечном итоге приводят к формированию физиологического ответа, обычно через изменение транскрипции генов. Цикл завершается гидролизом связанной с альфа-субъединицей ГТФ до ГДФ, что приводит к реассоциации α-субъединицы и βγ-гетеродимера и последующему связыванию гетеротримера G-белка с рецептором[3]. Длительность сигнала от G-белок-связанного рецептора зависит от стабильности ГТФ-связанной α-субъединицы, которая может регулироваться через белки RGS (англ. regulator of G protein signalling) или посттрансляционными модификациями[4]. Существует несколько изоформ каждой субъединицы, многие из которых имеют несколько сплайсинговых вариантов, что в сумме делает возможным образование тысяч различных G-белков. Определённая комбинация субъединиц в каждом гетеротримере G-белка влияет не только на то, с каким рецептором он может связаться, но также и на активируемые им эффекторы, что делает возможным направленный физиологический ответ для каждого типа стимулов[5][6]. На одной или нескольких субъединицах G-белка имеют липидные модификации, которые направляют их в определённую область на цитоплазматической мембране и вносят свой вклад в белок-белковое взаимодействие. Семейство состоит из субъединиц Gα, которые являются слабыми ГТФазами. G-белки обычно классифицируют в соответствии с их первичной структурой и функцией их субъединиц Gα, которые у млекопитающих подразделяются на несколько подтипов: Gsα, Gqα, Giα, трансдуцин, густдуцин и G12α; существует также несколько классов α-субъединиц растений и грибов. Каждая α-субъединица состоит из двух доменов: ГТФ-связывающего домена и спирального вставочного домена[7]. ГТФ-связывающий домен гомологичен Ras-подобным малым ГТФазам, и включает в себя два региона переключения I и II, которые осуществляют изменение конформации в момент активации G-белка. Переключатели представляют собой петли α-спиралей с конформацией, чувствительной к гуаниновым нуклеотидам. Спиральный вставочный домен расположен между ГТФ-связывающим доменом перед переключателем I и присущ только G-белкам. Он функционирует как изолятор для гуанинового нуклеотида от взаимодействия с ГТФ-связывающим доменом и абсолютно необходим для диссоциации нуклеотида.
Примечания
- Preininger A. M., Hamm H. E. G protein signaling: insights from new structures (англ.) // Sci. STKE : journal. — 2004. — February (vol. 2004, no. 218). — P. re3. — doi:10.1126/stke.2182004re3. — PMID 14762218.
- Roberts D. J., Waelbroeck M. G protein activation by G protein coupled receptors: ternary complex formation or catalyzed reaction? (англ.) // Biochem. Pharmacol. : journal. — 2004. — September (vol. 68, no. 5). — P. 799—806. — doi:10.1016/j.bcp.2004.05.044. — PMID 15294442.
- Svoboda P., Teisinger J., Novotný J., Bourová L., Drmota T., Hejnová L., Moravcová Z., Lisý V., Rudajev V., Stöhr J., Vokurková A., Svandová I., Durchánková D. Biochemistry of transmembrane signaling mediated by trimeric G proteins (англ.) // Physiol Res : journal. — 2004. — Vol. 53 Suppl 1. — P. S141—52. — PMID 15119945.
- Chen C. A., Manning D. R. Regulation of G proteins by covalent modification (англ.) // Oncogene. — 2001. — March (vol. 20, no. 13). — P. 1643—1652. — doi:10.1038/sj.onc.1204185. — PMID 11313912.
- Hildebrandt J. D. Role of subunit diversity in signaling by heterotrimeric G proteins (англ.) // Biochem. Pharmacol. : journal. — 1997. — August (vol. 54, no. 3). — P. 325—339. — doi:10.1016/S0006-2952(97)00269-4. — PMID 9278091.
- Albert P. R., Robillard L. G protein specificity: traffic direction required (англ.) // Cell. Signal. : journal. — 2002. — May (vol. 14, no. 5). — P. 407—418. — doi:10.1016/S0898-6568(01)00259-5. — PMID 11882385.
- (INTERPRO)