Liaodactylus
Liaodactylus (лат.) — род птерозавров-фильтраторов из семейства ктенохазматид, чьи окаменелые остатки обнаружены в юрской геологической формации Тяоцзишан в Китае. В род включают единственный вид Liaodactylus primus, описанный группой учёных под руководством Чан-Фу Жоу в 2017 году. Челюсти животного несли около 150 длинных, тонких, плотно расположенных зубов. Это самый ранний известный представитель семейства, а также первый птерозавр-фильтратор из формации Тяоцзишан. Поздние и более специализированные ктенохазматиды отличались от него более длинными мордами, меньшими по размеру окнами черепа и бо́льшим количеством зубов. Среди всех представителей семейства Liaodactylus наиболее тесно связан с европейским родом Ctenochasma.
| † Liaodactylus | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Череп голотипа | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Отряд: Подотряд: Надсемейство: Клада: † Ctenochasmatoidea Семейство: Род: † Liaodactylus |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Liaodactylus Zhou et al., 2017 | |||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
|
† Liaodactylus primus Zhou et al., 2017 |
|||||||||||||||||||||||||||||||||||||||||
| Геохронология 161,8—159,5 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
Открытие и название
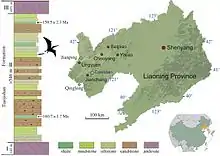
Известен только один образец Liaodactylus — голотип PMOL AP00031, который хранится в Палеонтологическом музее Ляонина. Он состоит из полного черепа с нижней челюстью и двух первых шейных позвонков. Образец происходит из пород, выходящих на поверхность примерно в 500 метрах к западу от деревни Дасишан, уезд Цзяньчан, провинция Ляонин (Китай). Эти породы относят к верхнеюрской формации Тяоцзишан, которую датируют оксфордским ярусом — от 161,8 до 159,5 млн лет назад[1][2][3].
Нового птерозавра назвала и описала команда палеонтологов под руководством Чан-Фу Жоу в 2017 году. Название рода сочетает в себе слово liao — сокращение от названия провинции Ляонин, и греч. δάκτυλος/daktylos — палец, типичное окончание названий птерозавров. Видовое название primus — первый, указывает на ранний возраст Liaodactylus относительно других ктенохазматид[1].
Описание
Liaodactylus является самым древним из известных ктенохазматид, на 8—10 миллионов лет старше титонских (152 миллиона лет) гнатозавра[4] и ктенохазмы. Кроме того, это первый оксфордский птеродактиль, который задокументирован с остатками черепа: наиболее базальный представитель группы Kryptodrakon, приблизительный современник Liaodactylus, представлен только неполным скелетом без черепа из верхней юры китайской формации Шишугоу[1].
Морда, челюсти и зубы
Голова Liaodactylus тонкая и длинная, с длиной черепа 13,3 сантиметра и длиной челюстей 11,7 сантиметра[1]. При этом длина челюстей составляет 49,1 % от общей длины черепа, а носо-предглазничное окно (наблюдаемое у всех представителей группы Monofenestrata[5]) — 31 %. У более продвинутых ктенохазматид носо-предглазничное окно намного меньше (например, всего 10—12 % у птеродаустро), а морда — длиннее (свыше 85 % длины черепа у птеродаустро)[6]. Симфиз Liaodactylus был короче такового у прочих ктенохазматид — 30,5 % длины челюсти. Морфологически стороны морды параллельны, в отличие от ложкообразного утолщения у гнатозавра. Бо́льшая часть нижней челюсти состоит в основном из зубной кости, а хорошо развитый ретроартикулярный отросток образован угловой костью, как у других представителей семейства[1].
Челюсти Liaodactylus содержат много длинных, игольчатых, выступающих наружу, плотно упакованных, равномерно расположенных зубов, в общей сложности до 152 зубов в обеих челюстях. Это больше, чем у Gnathosaurus (128—136), примерно столько же, сколько у Gegepterus (150)[7], но гораздо меньше, чем у ктенохазмы (200—552)[8] и птеродаустро (почти 1000)[6]. Зубы становятся короче к задней части челюстей, в конечном итоге превращаясь в короткие, похожие на колышки структуры в задней части зубного ряда. Задняя часть зубного ряда находится на одном уровне с передней третью носо-предглазничного окна, что необычно для ктенохазматид, у которых зубы перед носо-предглазничным окном исчезают. Первый зуб в передней части каждой челюсти также несколько укорочен и составляет только половину длины второго. При сомкнутых челюстях зубы образуют структуру, похожую на расчёску, которая служила для фильтрации воды[1][9].
Черепная коробка и позвонки
Верхняя часть черепной коробки состоит из сильно сросшихся лобной и теменной костей, которые резко изгибаются около затылка, образуя закруглённый свод черепа. Базисфеноид в задней части черепа короткий, широкий и плоский, с широкой U-образной выемкой на переднем конце. По обе стороны от выемки лежит базисфеноидный отросток, который соединяется с клиновидной и квадратной костями. Сзади базисфеноид становится у́же и перекрывается базиокципитальной костью. Затылочный мыщелок, который соединяет череп с позвонками, имеет сферическую форму, а отверстие для спинного мозга, foramen magnum, скрыто, когда позвонки сочленены с черепом. Имеется шероховатый гребень, идущий вдоль средней линии базиокципитальной кости вперёд от затылочного мыщелка[1].
Первый и второй шейные позвонки слиты, хотя и с видимым швом между ними. В передней части первого шейного позвонка имеется маленький рудимент (про-атлант) — он симметричный, уплощённый и имеет выступ в передней части. Сферическому затылочному мыщелку соответствует чашеобразный сустав атланта. Атлант несёт заметную невральную дугу, но невральная дуга на эпистрофее не видна. В задней части эпистрофея, на невральной дуге, расположен постзигапофиз, который должен соединяться со следующим шейным позвонком[1].
Систематика
На основе проведённого филогенетического анализа Жоу с соавторами отнесли Liaodactylus к семейству ктенохазматид. С этим семейством птерозавра сближают такие черты, как относительная высота черепа на уровне чешуйчатой кости, высота черепа на уровне челюстного сустава и длина морды на уровне носо-предглазничного окна. Кроме того, на основе морфологических признаков, череп птерозавра несёт ретроартикулярный отросток, направленный вниз и назад. В пределах семейства Liaodactylus оказался тесно связан с ктенохазмой, с которой он разделяет отношение длины челюсти к черепу и наклона зубов в стороны. Кладограмма, построенная в результате анализа, изображена ниже[1][5]:
| Pterodactyloidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеоэкология
Liaodactylus известен из геологической формации Тяоцзишан, которая, вместе с более древней формацией Хайфангоу образуют биоту Янляо — разнообразную группу хорошо сохранившихся юрских животных, содержащую по меньшей мере 40 видов. Здесь был обнаружен ряд других птерозавров, включая рамфоринхид Jianchangnathus robustus и Qinglongopterus guoi; анурогнатид Dendrorhynchoides mutoudengensis и Jeholopterus ningchengensis; представителей подгруппы Darwinoptera Pterorhynchus wellnhoferi и семейства Wukongopteridae Darwinopterus modularis и Wukongopterus lii; а также, возможно, Archaeoistiodactylus linglongtaensis[10]. При этом Liaodactylus является первым птерозавром-фильтратором биоты Янляо[1]. Динозавры представлены скансориоптеригидами Epidendrosaurus ninchengensis, Epidexipteryx hui и Yi qi и представителями клады Paraves, такими как Anchiornis huxleyi, Pedopenna daohugouensis и Xiaotingia zhengi[11][12].
Неархозавровые формы также присутствуют в биоте Янляо. Древняя экосистема включала млекопитающих[13][14], ящериц, в том числе, «Yabeinosaurus» youngi[15]; не описанного крокодиломорфа; саламандр[16] и рыб, в том числе вид Liaosteus hongi и представителя семейства Ptycholepidae[11][17].
С точки зрения окружающей среды, формация Тяоцзишан представляла собой лесистую местность с хвойными деревьями, саговниками и папоротниками. Найденные здесь виды ископаемых растений включают хвощ Neocalamites carcinoides, N. narthosi и Equisetum sp.; древовидные папоротники Coniopteris hymenophyloides и C. margaretae; саговник Zamites gigas; загадочные голосеменные Czekanowskia ketova, C. ridiga и Phoenicopsis speciosa; гинкго Sphenobaiera kazachstanica; кипарис Protaxodioxylon jianchangense и неопределённое хвойное дерево Xenoxylon peidense. В совокупности эти растения указывают на прохладный, умеренный, влажный и сезонный климат со средней температурой ниже 15 °C[17]. Это согласуется с тенденцией глобального похолодания примерно в это время[18], но контрастирует с тёплым, сухим климатом более ранней формации Хайфангоу[19].
Примечания
- Zhou C.-F., Gao K.-Q., Yi H., Xue J., Li Q., Fox R. C. Earliest filter-feeding pterosaur from the Jurassic of China and ecological evolution of Pterodactyloidea (англ.) // Royal Society Open Science. — 2017. — Vol. 4, no. 2. — P. 160672. — doi:10.1098/rsos.160672.
- Chang S.-C., Zhang H., Renne P. R., Fang Y. High-precision 40Ar/39Ar age constraints on the basal Lanqi Formation and its implications for the origin of angiosperm plants (англ.) // Earth and Planetary Science Letters. — 2009. — Vol. 279, no. 3—4. — P. 212—221. — doi:10.1016/j.epsl.2008.12.045.
- Chang S.-C., Zhang H., Hemming S. R., Mesko G. T., Fang Y. 40Ar/39Ar age constraints on the Haifanggou and Lanqi formations: When did the first flowers bloom? (англ.) // Geological Society, London, Special Publications. — 2013. — Vol. 378. — P. 277—284. — doi:10.1144/SP378.1.
- Татаринов Л. П. Глава VI. Птерозавры (Pterosauria), или летающие ящеры // Очерки по эволюции рептилий. Архозавры и зверообразные. — М. : ГЕОС, 2009. — С. 175—195. — 377 с. : ил. — (Труды ПИН РАН ; т. 291). — 600 экз. — ISBN 978-5-89118-461-9.
- Andres B., Clark J., Xu X. The Earliest Pterodactyloid and the Origin of the Group (англ.) // Current Biology. — 2014. — Vol. 24, no. 9. — P. 1011—1016. — doi:10.1016/j.cub.2014.03.030. (недоступная ссылка)
- Chiappe L. M., Kellner A. W. A., Rivarola D., Davila S., Fox M. Cranial Morphology of Pterodaustro guinazui (Pterosauria: Pterodactyloidea) from the Lower Cretaceous of Argentina (англ.) // Natural History Museum of Los Angeles Contributions in Science. — 2000. — Vol. 483. — P. 1—19.
- Wang X., Kellner A. W. A., Zhou Z., de Almeida Campos D. A new pterosaur (Ctenochasmatidae, Archaeopterodactyloidea) from the Lower Cretaceous Yixian Formation of China (англ.) // Cretaceous Research. — 2007. — Vol. 28, no. 2. — P. 245–260. — doi:10.1016/j.cretres.2006.08.004.
- Bennett S. C. A review of the pterosaur Ctenochasma: taxonomy and ontogeny (англ.) // Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. — 2007. — Vol. 245, no. 1. — P. 23—31. — doi:10.1127/0077-7749/2007/0245-0023.
- Witton M. P. 19. Ctenochsmatiodea // Pterosaurs: Natural History, Evolution, Anatomy. — Princeton: Princeton University Press, 2013. — 291 с. — ISBN 978-0-691-15061-1.
- Witton M. P. Were early pterosaurs inept terrestrial locomotors? (англ.) // PeerJ. — 2015. — Vol. 3. — P. e1018. — doi:10.7717/peerj.1018.
- Zhou Z.-H., Wang Y. Vertebrate assemblages of the Jurassic Yanliao Biota and the Early Cretaceous Jehol Biota: Comparisons and implications (англ.) // Paleoworld. — 2017. — Vol. 26, no. 2. — P. 241. — doi:10.1016/j.palwor.2017.01.002.
- Sullivan C., Wang Y., Hone D. W. E., Wang Y., Xu X., Zhang F. The vertebrates of the Jurassic Daohugou Biota of northeastern China (англ.) // Journal of Vertebrate Paleontology. — 2013. — Vol. 34, no. 2. — P. 243—280. — doi:10.1080/02724634.2013.787316.
- Meng J. Mesozoic mammals of China: implications for phylogeny and early evolution of mammals (англ.) // National Science Review. — 2014. — Vol. 1, no. 4. — P. 521—542. — doi:10.1093/nsr/nwu070.
- Luo Z.-X., Meng Q.-J., Ji Q., Liu D., Zhang Y.-G., Neander A. I. Evolutionary development in basal mammaliaforms as revealed by a docodontan (англ.) // Science. — 2015. — Vol. 347, no. 6223. — P. 760—764. — doi:10.1126/science.1260880.
- Evans S. E., Wang Y. New material of the Early Cretaceous lizard Yabeinosaurus from China (англ.) // Cretaceous Research. — 2012. — Vol. 34. — P. 48–60. — doi:10.1016/j.cretres.2011.10.004.
- Gao K.-Q. Chen J., Jia J. Taxonomic diversity, stratigraphic range, and exceptional preservation of Juro-Cretaceous salamanders from northern China (англ.) // Canadian Journal of Earth Sciences. — 2013. — Vol. 50, no. 3. — P. 255—267. — doi:10.1139/e2012-039.
- Tian N., Xie A.-W., Wang Y.-D., Jiang Z.-K., Li L.-Q., Yin Y.-L., Zhu Z.-P., Wang J.-J. New records of Jurassic petrified wood in Jianchang of western Liaoning, China and their palaeoclimate implications (англ.) // Science China Earth Sciences. — 2015. — Vol. 58, no. 12. — P. 2154–2164. — doi:10.1007/s11430-015-5208-1.
- Dromart G., Garcia J.-P., Picard S., Atrops F., Lecuyer C., Sheppard S. M. F. Ice age at the Middle–Late Jurassic transition? (англ.) // Earth and Planetary Science Letters. — 2003. — Vol. 213, no. 3. — P. 205—220. — doi:10.1016/S0012-821X(03)00287-5.
- Na Y., Manchester S.R., Sun C., Zhang S. The Middle Jurassic palynology of the Daohugou area, Inner Mongolia, China, and its implications for palaeobiology and palaeogeography (англ.) // Palynology. — 2015. — Vol. 39, no. 2. — P. 270. — doi:10.1080/01916122.2014.961664.