Пауссины
Пауссины (лат. Paussinae) — подсемейство жуков из семейства жужелиц. В России — 1 вид.
| Пауссины | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
.jpg.webp) Cerapterus pilipennis | ||||||||||
| Научная классификация | ||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Без ранга: Тип: Подтип: Надкласс: Класс: Подкласс: Инфракласс: Надотряд: Отряд: Подотряд: Надсемейство: Семейство: Подсемейство: Пауссины |
||||||||||
| Международное научное название | ||||||||||
| Paussinae Latreille, 1807 | ||||||||||
| Трибы | ||||||||||
|
||||||||||
| ||||||||||
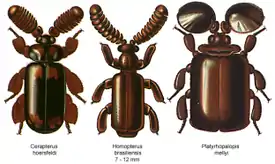





Описание
Жуки от мелких до средних размеров; в длину достигают от 2,8 до 22 мм. Окраска жуков преимущественно коричневатая, варьирует от почти чёрной до бледноватой коричнево-красной. Очень явным отличительным признаком подсемейства являются их большие в диаметре усики. Жуки триб Metriini и Ozaenini имеют форму тела более типичную для жужелиц, , но некоторые роды демонстрируют признаки, которые напоминают мирмекофильные адаптации других жуков, такие как толстые усики, редукция тактильных щетинок или уплощенное тело и ноги. Специализированную форму тела имеют трибы Protopaussini и Paussini, у которых отражены две стратегии адаптации к мирмекофильному образу жизни: они либо узкие и тонкие, или сплющенные и широкие. Ни один из этих жуков не похож по форме на муравьёв и не является мирмекоморфным. Для многих видов трибы Paussini характерны увеличенные усики[1].
У более специализированных видов ротовой аппарат жука имеет форму ложки, чтобы легко получать капли пищи из ротового аппарата муравья. Более примитивные виды больше полагаются на грубую силу и защищаются жестким хитиновым покровом и бомбардируют нападающих муравьёв (или человека) газом, содержащим хинон, выделяемый специальными железами. Газ испаряется со взрывом, высвобождая облако пара, так ка это происходит у других жуков-бомбардиров[2].
Экология и местообитания
Почти все представители пауссин — мирмекофилы, и живут ассоциациями с муравьями. В связи с этим и произошли английские названия «ants' guest beetles» — «жуки-гости муравьёв», и «ants' nest beetles» — «муравейниковые жуки»[2]. Среди муравьёв-хозяев преобладают представители подсемейств мирмицины и формицины, редко долиходерины, Ponerinae и Ectatomminae[1]. Представители неспециализированной трибы Metriini внешне похожи на обычных жужелиц, питаются многоножками; личинки свободноживущие, не связанные с муравьями[1].
Пауссины выделяют летучие вещества (феромоны) из подкожных желёз, которые действуют умиротворяюще на муравьёв и подавляют их обычную агрессию против «злоумышленников». Взамен, жуки и их личинки получают пищу муравьёв, а также личинки жуков не прочь полакомиться и личинками самих муравьёв[2]. Хотя пауссины принимаются и интегрируются в колонию муравьёв-хозяев, они наносят некоторый ущерб колонии, охотясь на муравьёв и их потомство. Такой частичный симбиоз, при котором один партнер живёт за счёт другого — в данном случае пауссины, поражающие муравьиную колонию, — называется паразитизмом (Hölldobler and Wilson, 1990). Представители специализированной трибы Paussini, однако, выделяют муравьям особый секрет своих желёз, который Эрих Васманн (1903) считал достаточной компенсацией за потерю потомства, чтобы сделать отношения взаимовыгодными[1].
Представители триб Metriini и Ozaenini встречаются под камнями, поваленными деревьями и корой, а также в гнилой древесине, опавших листьях и перегное. Paussini обычно встречаются в приземной зоне растительности и в основном собирались из подземных муравейников. Однако на присутствие пауссин на деревьях указывают спорадические наблюдения, так что можно предположить, что Paussini поселяются в гнезде своих муравьёв-хозяев, независимо от его местонахождения. Помимо муравейников, самцы Paussini отлавливаются световыми ловушками, тогда как самок таким способом ловят редко. Различные другие методы отлова дают небольшое количество Paussini, которые, однако, часто представляют виды, обычно не регистрируемые в световых ловушках[1].
Активность пауссин (нахождение вне муравейников) зависит от сезонных климатических условий. Прямыми триггерами могут быть влажность, температура или продолжительность дня, которые также влияют на колонии муравьёв и, таким образом, косвенно влияют на активность пауссин. Внетропические Metriini активны в весенний и летний периоды в Калифорнии. В областях с зимними дождями, субтропическим или тропическим сезонным климатом они редки, Paussini там наблюдаются в сухой сезон, тогда как они становятся более обильными при грозовой атмосфере и дожде. Во внутренних тропиках Африки Paussini активны в течение всего года, но с пиками численности. Что касается циркадной активности, то большинство пауссин ведут сумеречный или ночной образ жизни, хотя существуют и дневные виды[1][3][4].
Распространение
Пауссины встречаются на всех континентах, кроме Антарктиды, но их основное распространение ограничено субтропическими и тропическими областями с наибольшим разнообразием в палеотропике. Неарктические Metriini являются внетропическими, в то время как несколько представителей Ozaenini, Protopaussini и Paussini достигают самых южных частей Неарктики и Палеарктики. Окаменелости янтаря из европейской части Балтийского моря и Северного моря, а также окаменелости из озерных отложений из Германии свидетельствуют о том, что Paussini когда-то были распространены в районах, где они вымерли сегодня[1][5][6].
Палеонтология
Ископаемые находки пауссин известны из балтийского и других видов янтарей, возрастом около 50 млн лет (Eohomopterus poinari; эоцен-миоцен; 45-15 млн лет)[5] и из бирманского янтаря мелового периода (Kryzhanovskiana olegi, около 100 млн лет)[7].
Доминиканские (остров Гаити) и мексиканские янтари (Чьяпас) возникли в сходных условиях и документируют раннетретичное распространение на южной окраине Западной Лавразии. Ареалы некоторых видов Paussus соответствуют определенным зонам растительности, что указывает на связь с крупномасштабными климатическими и экологическими условиями. Более того, на экорегиональную, а не связанную с муравьями-хозяином зависимость распространения жуков указывает северный предел распространения Paussus favieri на юге Франции, который расположен южнее, чем предел распространения его основного муравья-хозяина[1][8][9][10].
Систематика
Филогения
Эволюция пауссин шла в направлении от свободноживущего хищного образа жизни к мирмекофилии. Морфологические признаки, такие как трихомы и специфические железы, четко классифицируют наиболее специализированную трибу Paussini как характерных мирмекофилов, а их ротовой аппарат узко специализирован для добычи муравьёв[1]. По результатам анализа морфологии личинок Paussinae выдвинуты две рабочие гипотезы (Giulio, 2003) относительно филогении этой группы жуков. По одной гипотезе Metriini являются сестринской группой для Ozaenini + Paussini, где Ozaenini является парафилетической группой, а род Physea рассматривается в качестве сестринской группы к трибе Paussini. Согласно второй гипотезе Metriini являются сестринской группой для ветвистоусых Ozaenini, а Paussini сестринской для клады Metriini + Ozaenini[11].
Схема филогении трибы пауссин
└─o Carabidae
└─o Paussinae
├─o Metriini
└─o
├─o Protopaussini
└─o
├─o Ozaenini
└─o Paussini
Классификация
Более 750 видов в 4 трибах[12]. Среди крупнейших родов таксоны Paussus (329 видов), Arthropterus (66), Pachyteles (49)[1][13][14][15].
- триба Metriini LeConte, 1853
- Metrius Eschscholtz, 1829
- Sinometrius Wrase & J.Schmidt, 2006
- триба Ozaenini Hope, 1838
- Anentmetus Andrewes, 1924
- Anentmetus pluto Andrewes, 1924
- Anentmetus spissicornis Fairmaire, 1888
- Entomoantyx Ball et McCleve, 1990
- Entomoantyx cyanipennis Chaudoir, 1852
- Falcopaussus
- Filicerozaena Deuve, 2001
- Filicerozaena moreti Deuve, 2001
- Filicerozaena tapiai
- Inflatozaena Deuve, 2001
- Inflatozaena inflata Bates, 1884
- Itamus Loew, 1849
- Itamus castaneus Schmidt-Goebel, 1846
- Itamus cavicola Moore, 1978
- Itamus dentatus Andrewes, 1919
- Itamus kaszabi Jedlicka, 1968
- Mystropomus Chaudoir, 1848 LinkOut
- Mystropomus regularis Banninger, 1940
- Mystropomus subcostatus Chaudoir, 1848
- Ozaena Olivier, 1812
- Ozaena boucheri Deuve, 2001
- Ozaena convexa Banninger, 1927
- Ozaena dentipes Olivier, 1812
- Ozaena ecuadorica Banninger, 1949
- Ozaena elevata Banninger, 1956
- Ozaena grossa Banninger, 1927
- Ozaena lemoulti Banninger, 1932
- Ozaena linearis Banninger, 1927
- Ozaena manu Ball & Shpeley, 1990
- Ozaena martinezi Ogueta, 1965
- Ozaena maxi Ball & Shpeley, 1990
- Pachyteles
- Pachyteles (Goniotropis) Gray, 1832
- Pachyteles (Pachyteles) Perty, 1830
- Pachyteles (Tropopsis) Solier, 1849
- Physea Brulle, 1834
- Platycerozaena Banninger, 1927
- Platycerozaena bordoni Ogueta, 1965
- Platycerozaena brevicornis Bates, 1874
- Platycerozaena magna Bates, 1874
- Platycerozaena panamensis Banninger, 1949
- Proozaena Deuve, 2001
- Proozaena parallela Chaudoir, 1848
- Pseudozaena Laporte, 1834
- Pseudozaena orientalis Klug, 1834
- Pseudozaena tricostata Montrouzier, 1855
- Serratozaena Deuve, 2001
- Serratozaena paraphysea Deuve, 2001
- Sphaerostylus Chaudoir, 1848
- Sphaerostylus (Afrozaena) Jeannel, 1946
- Sphaerostylus (Ozaeniella) Basilewsky, 1976
- Sphaerostylus (Sphaerostylus) Chaudoir, 1848
- Tachypeles Deuve, 2001
- Tachypeles lecordieri Deuve, 2001
- Anentmetus Andrewes, 1924
- триба Paussini Latreille, 1807
- подтриба Arthropterina Nagel, 1987
- Arthropterus MacLeay, 1838
- Megalopaussus Lea, 1906
- подтриба Carabidomemnina Wasmann, 1928
- подтриба Cerapterina Billberg, 1820
- Cerapterus Swederus, 1788
- подтриба Heteropaussina Janssens, 1953
- Heteropaussus J.Thomson, 1860
- подтриба Homopterina Wasmann, 1920
- подтриба Paussina Latreille, 1807
- Ceratoderus Westwood, 1841
- Ceratoderus jendeki
- Granulopaussus H.Kolbe, 1938
- Hylopaussus Luna de Carvalho, 1989
- Hylotorus Dalman, 1823
- Leleupaussus Luna de Carvalho, 1962
- Melanospilus Westwood, 1845
- Paussomorphus Raffray, 1885
- Paussus Linne, 1775
- Ceratoderus Westwood, 1841
- подтриба Platyrhopalina
- Euplatyrhopalus Desneux, 1905
- Lebioderus Westwood, 1838
- Platyrhopalus Westwood, 1833
- Platyrhopalopsis Desneux, 1905
- Pterorhopalus Maruyama, 2011
- подтриба Pentaplatarthrina Jeannel, 1946
- Hexaplatarthrus Jeannel, 1955
- Pentaplatarthrus Westwood, 1833
- подтриба Arthropterina Nagel, 1987
- триба Protopaussini Gestro, 1892
- Protopaussus Gestro, 1892
Примечания
- Stefanie F. Geiselhardt, Klaus Peschke, Peter Nagel. A review of myrmecophily in ant nest beetles (Coleoptera: Carabidae: Paussinae): linking early observations with recent findings. (англ.) // Naturwissenschaften : Журнал. — 2007. — Vol. 94, no. 11. — P. 871—894. — doi:10.1007/s00114-007-0271-x.
- Subfamily: Paussinae (ants' guest beetles, ants' nest beetles). biodiversityexplorer.info
- Kistner D. H. (1982) The social insects’bestiary. In: Hermann HR (ed) Social insects, vol. 3. Academic, New York, pp 1–244
- Batelka J (2000) A contribution to the knowledge of the bionomics and distribution of two species of the subfamily Paussinae (Coleoptera: Carabidae) in Morocco and Tunisia. Klapalekiana 36:217–223
- Nagel P. 1987. Fossil ant nest beetles (Coleoptera, Carabidae, Paussinae). «Entomologische Arbeiten aus dem Museum G. Frey». 35/36, ss. 137—170 (1987). (недоступная ссылка)(8,5 MB)
- Wasmann E. (1929) Die Paussiden des baltischen Bernsteins und die Stammesgeschichte der Paussiden. 270. Beitrag zur Kenntnis der Myrmecophilen. Bernsteinforschungen 1:1–110
- Boris M. Kataev, Alexander G. Kirejtshuk, Andranik R. Manukyan, Boris A. Anokhin. Kryzhanovskiana olegi gen. et sp. nov., a remarkable eyeless representative of the tribe Metriini (Coleoptera: Carabidae: Paussinae) from Upper Cretaceous amber of northern Myanmar (англ.) // Cretaceous Research : Журнал. — Elsevier Ltd., 2019. — Vol. 103, no. 104168. — P. 1—13. — ISSN 0195-6671. — doi:10.1016/j.cretres.2019.06.014.
- Nagel P. (1997) New fossil paussids from Dominican amber with notes on the phylogenetic systematics of the paussine complex (Coleoptera: Carabidae). Syst Entomol 22:345–362
- Solorzano Kraemer M. (2006) The first fossil paussine (Coleoptera: Carabidae) from Mexican amber. Palaontol Z 80:107–111
- Le Masne G. (1961) Observations sur le comportement de Paussus favieri Fairm., hôte de la fourmiPheidole pallidulaNyl. Ann Fac Sci Marseille 31:111–130
- Di Giulio A., Fattorini S., Kaupp A., Vigna Taglianti A., Nagel P. 2003. Review of competing hypotheses of phylogenetic relationships of Paussinae (Coleoptera: Carabidae) based on larval characters. «Systematic Entomology». 28. 4, pp. 509—537 (2003). doi:10.1046/j.1365-3113.2003.00227.x
- Nagel P. 2003. New fossil paussids from Dominican amber with notes on the phylogenetic systematics of the paussine complex (Coleoptera: Carabidae). «Systematic Entomology». 22. 4, ss. 345—362 (2003). doi:10.1046/j.1365-3113.1997.d01-51.x
- Nagel P. (2006) Ant nest beetles of the Carnegie Museum (Coleoptera, Carabidae, Paussinae, Paussini). Ann Carnegie Mus 75:181-202
- Deuve T. (2004) Nouveaux Ozaenini Néotropicaux (Coleoptera, Paussidae). Rev Fr Entomol NS 26:117-130
- Deuve T. (2005) Nouveaux Ozaeninae (Eustrini et Ozaenini) de la Région Néotropicale et du Népal (Coleoptera, Caraboidea, Paussidae). Rev Fr Entomol NS 27:103-116
