Моторная кора
Моторная кора, также известная как двигательная кора (англ. Motor cortex) — области коры больших полушарий, отвечающих за планирование, контроль и выполнение произвольных движений.
| Моторная кора | |
|---|---|
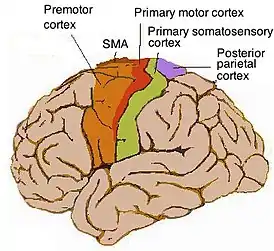 Топография моторной коры головного мозга человека | |
| Часть | Кора больших полушарий |
| Каталоги | |
Традиционно моторной корой считается область в лобной доле, расположенная в задней части прецентральной извилины непосредственно перед центральной бороздой.
Компоненты моторной коры
Моторная кора состоит из трёх областей:
1. Первичная моторная кора (англ. Primary motor cortex) соответствует полю 4 по Бродману, располагается кпереди от центральной борозды в передней части околоцентральной дольки. Главный корковый центр произвольных движений. Получает афферентную информацию от мозжечка (через таламус), от премоторной коры и дополнительной моторной области, сенсорных областей коры. Собственно произвольное движение происходит, когда потенциал действия от нейронов первичной моторной коры по аксонам нисходящих путей достигает сегментов спинного мозга и α-мотонейроны передних рогов вызовут движения. То же самое происходит и при стимуляции первичной моторной коры электродом во время нейрохирургической операции[1].
2. Премоторная кора (англ. Premotor cortex) соответствует латеральной части поля 6 по Бродману, располагается кпереди (латеральнее) от прецентральной борозды в задней части трёх горизонтальных лобных извилин (верхней, средней и нижней)[1]. Отвечает за некоторые элементы моторного контроля, включая планирование точных серий сокращений мышц, регуляцию положения тела в пространстве (осанки) и т.д. Премоторная кора располагается перед первичной моторной корой.
3. Дополнительная моторная область (англ. Supplementary motor area, SMA) соответствует медиальной части поля 6 по Бродману, располагается на медиальной поверхности полушарий. Отвечает за планирование движений, координацию двух частей тела. Стимуляция данной области приводит к одновременному сжиманию кистей обеих рук, что, вероятно, является рудиментарной функцией кистей, необходимой для лазания[1].
Другие области:
- Задняя теменная кора иногда также относится к числу областей, отвечающих за моторную активность, хотя это ассоциативная, а не двигательная кора. В данной области аккумулируется информация от разных сенсорных систем, после чего она преобразуется в двигательные команды. Задняя теменная кора задействована в некоторых механизмах планирования движений, в дополнение к многим другим функциям, которые не связаны с моторным контролем.
- Первичная соматосенсорная кора, особенно зона 3a, расположенная в постцентральной извилине, иногда считается функциональной частью нейросети двигательного контроля.
Другие области мозга, расположенные вне коры больших полушарий, также оказывают большое влияние на двигательные функции. К ним относятся мозжечок, базальные ганглии, педункулопонтийное тегментальное ядро и красное ядро, а также некоторые другие субкортикальные моторные ядра.
Эволюция моторной коры
Млекопитающие эволюционировали из зверообразных рептилий около 200 млн лет назад[2]. У ранних млекопитающих появились новые мозговые функции, отвечающие тем нишам, которые эти животные заняли в природе[3]. У них появилась соматомоторная кора, в которой обрабатывалась как соматосенсорная, так и двигательная информация. Это позволило появиться только простым моторным навыкам, например, передвижению на четырёх ногах, убеганию от хищников или погоне за жертвой. У плацентарных млекопитающих появилась отдельная моторная кора, это произошло около 100 млн лет назад[2]. Так как масса нервной ткани, отвечающей за определённую функцию, соответствует объёму информации, который обрабатывается при выполнении данной функции[3], развитие обособленной моторной коры давало преимущество плацентарным млекопитающим, а их двигательные навыки становились более сложными, чем у их предков. В дальнейшем моторная кора сыграла важную роль в адаптации приматов к древесному образу жизни.
Появление моторной коры и её усложнение (а также появление противопоставленного большого пальца руки и стереоскопического зрения) поддерживалось естественным отбором приматов, передвигающихся на ветках деревьев (Cartmill, 1974; Silcox, 2007). В результате такого отбора возникло непропорциональное соматотопическое представительство рук и ног, которые были важны для хватания веток и передвижений по деревьям (Nambu, 2011; Pons et al., 1985; Gentilucci et al., 1988).
Примечания
- Ерофеев Н.П. Физиология центральной нервной системы. Учебное пособие. — Москва: СпецЛит, 2017. — С. 148-149. — ISBN 78-5-299-00841-8.
- Kaas, J.H. Evolution of somatosensory and motor cortex in primates (англ.) // The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology : journal. — 2004. — Vol. 281, no. 1. — P. 1148—1156. — doi:10.1002/ar.a.20120. — PMID 15470673.
- Jerison, Harry. Evolution of the Brain and Intelligence. — Academic Press Inc., 1973.