Митотический кроссинговер
Митоти́ческий кроссинго́вер — тип генетической рекомбинации, который может проходить в соматических клетках при митотических делениях как у организмов, обладающих полом, так и бесполых организмов (например, некоторых одноклеточных грибов, у которых не известен половой процесс). В случае бесполых организмов митотическая рекомбинация является единственным ключом к пониманию сцепления генов, так как у таких организмов это единственный способ генетической рекомбинации[1]. Кроме того, митотическая рекомбинация может привести к мозаичной экспрессии рецессивных признаков у гетерозиготной особи. Такая экспрессия имеет важное значение в онкогенезе, она также позволяет изучать летальные рецессивные мутации[1][2].
Открытие
Открытие митотической рекомбинации было совершено Куртом Штерном в 1936 году в результате исследования дрозофил, являвшихся гетерозиготами по двум генам, находящихся на одной Х-хромосоме и определяющих легко регистрируемые признаки: цвет тела и форму щетинок. Рецессивный аллель первого гена y давал жёлтую окраску тела, рецессивный аллель sn (singed) — опалённые щетинки. Исследуемые дигетерозиготы с генотипом y+//+sn имели дикий фенотип по обоим признакам, однако изредка на теле некоторых мух появлялись двойные пятна. У таких пятен одна половина была жёлтая и с нормальными щетинками (доминантный признак), а другая — нормального серого цвета (доминантный признак), но с опалёнными щетинками. Подобные пятна были впервые замечены ещё в 1925 году, но лишь позднее Курт Штерн объяснил их появление митотическим кроссинговером на стадии 4-х хроматид[3][4].
Общие сведения
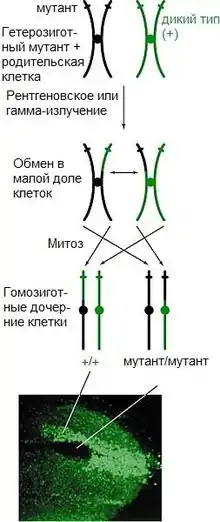
Митотическая рекомбинация может происходить в любом локусе, однако её результат виден лишь тогда, когда особь гетерозиготна по этим локусам. В результате митотического кроссинговера при расхождении хромосом в половине случаев должны образовываться двойные пятна, так как в двух случаях из четырёх одна из пары хроматид будет нормальной, а другая — рекомбинантной.
Иными словами, фенотип дочерней клетки зависит от того, как разойдутся хромосомы по метафазной пластинке. Если две рекомбинантные хроматиды, содержащие различные аллели, встанут на пластинке друг против друга, то дочерняя клетка будет гетерозиготной и с нормальным фенотипом, несмотря на рекомбинацию (так как рекомбинантные участки как бы взаимно компенсируют друг друга). Если же против рекомбинантной хроматиды станет нормальная, то клетка будет гомозиготной по рекомбинантному локусу. Результатом этого будет появление двойного пятна, где у некоторых клеток будет проявляться гомозиготный рецессивный фенотип, а у других — гомозиготный доминантный (дикий) фенотип. Если такие дочерние клетки продолжат размножаться и делиться, двойные пятна будут расти, приводя к появлению новых фенотипов.
Митотическая рекомбинация происходит в течение интерфазы. Было высказано предположение, что рекомбинация происходит в течение G1-фазы, когда хромосома ещё не удвоена[5]. В более недавнее время было предположено, что митотический кроссинговер происходит вследствие репарации путём гомологичной рекомбинации двунитевых повреждений ДНК, а сам процесс идёт в G1-фазе, однако этому противоречит то, что репарация путём гомологичной рекомбинации происходит в основном после репликации[6].
Молекулярные механизмы
Молекулярные механизмы, обеспечивающие митотический кроссинговер, схожи с участвующими в мейотическом кроссинговере. К ним относятся: образование гетеродуплексов, гомологичная рекомбинация, инициируемая двунитевыми разрывами ДНК, образование полухиазм и структур Холлидея. Возможно также, хотя и с значительно более низкой вероятностью, участие механизмов репарации повреждений ДНК путём негомологичной рекомбинации[6][7][8].
Хромосомные механизмы
Существует несколько теорий, объясняющих, как происходит митотический кроссинговер. В простейшей модели кроссинговера две хроматиды перекрываются или сближаются в конститутивных ломких сайтах, которые предрасположены к разрывам. При возникновении двуцепочечного разрыва в ломком сайте[9], он устраняется путём репарации на основе одной из двух сестринских хроматид другой хромосомы. Это позволяет двум хромосомам обмениваться участками. В другой модели две перекрывающиеся хромосомы образуют структуры Холлидея на общем повторяющемся участке. Потом эта крестообразная структура рассекается так, что одна хромосома обменивается участком одной из хроматид с участком хроматиды другой гомологичной хромосомы. В обеих моделях митотический кроссинговер является лишь из одним из возможных исходов.
С другой стороны, кроссинговер может происходить в ходе репарации ДНК[10], если хромосома послужила шаблоном для репарации гомологичной повреждённой хромосомы.
Преимущества и недостатки
Митотический кроссинговер наблюдали у D. melanogaster, у некоторых размножающихся бесполым путём грибов и в соматических клетках человека и мыши. В последнем случае митотический кроссинговер может приводить к появлению клеток, экспрессирующих рецессивные проонкогенные мутации, предрасполагая к развитию рака. С другой стороны, клетка может стать и гомозиготным мутантом по гену-супрессору опухолевого роста, что приведёт к тому же самому результату[2]. Например, синдром Блума вызывается мутацией RecQ-хеликазы, которая играет роль в репликации и репарации ДНК. У мышей эта мутация ведёт к повышению частоты митотической рекомбинации, что в свою очередь повышает частоту развития опухолей[11].
В то же время митотический кроссинговер может иметь и положительный эффект для организма, у которого доминантные аллели в гомозиготном состоянии более функциональны, чем в гетерозиготном[2].
Для экспериментов с геномами модельных организмов, например, дрозофилы, митотический кроссинговер можно вызвать искусственно рентгеновским излучением и FLP-FRT-рекомбинацией[12].
Примечания
- Hartel, Daniel L. and Maryellen Ruvolo. Genetics: Analysis of Genetics and Genomes (англ.). — Burlington: Jones & Bartlett, 2012.
- Tischfield, Jay A. Loss of Heterozygosity, or: How I learned to Stop Worrying and Love Mitotic Recombination (англ.) // American Journal of Human Genetics : journal. — 1997. — November (vol. 61, no. 5). — P. 995—999.
- Stern, Curt. Somatic Crossing Over and Segregation in Drosophila Melanogaster (англ.) // Genetics : journal. — 1936. — Vol. 21, no. 6. — P. 625—730.
- Инге-Вечтомов С.Г. Генетика с основами селекции: учебник для студентов высших учебных заведений / С. Г. Инге-Вечтомов. — СПб.: Изд-во Н-Л, 2010. — С. 193—194. — 720 с. — ISBN 978-5-94869-105-3.
- Esposito, Michael S. Evidence that Spontaneous Mitotic Recombination Occurs at the Two-Strand Stage (англ.) // Proceedings of the National Academy of Sciences of the United States of America : journal. — 1978. — September (vol. 75, no. 9). — P. 4436—4440.
- LaFave, MC; J Sekelsky. Mitotic Recombination: Why? When? How? Where? (англ.) // PLoS Genet : journal. — 2009. — Vol. 5, no. 3. — doi:10.1371/journal.pgen.1000411.
- Helleday, Thomas. Pathways for Mitotic Homologous Recombination in Mammalian Cells (англ.) // Mutation Research : journal. — Elsevier, 2003. — Vol. 532, no. 1—2. — P. 103—115.
- Pâques, Frédéric; James E. Haber. Multiple Pathways of Recombination Induced by Double-Strand Breaks in Saccharomyces cerevisiae (англ.) // Microbiology and Molecular Biology Reviews : journal. — American Society for Microbiology, 1999. — Vol. 63, no. 2. — P. 349—404.
- Helleday, T. Double-Strand Break Repair via Double Holliday Junctions (Szostak Model). Animation. MIT. Дата обращения: 1 марта 2013. Архивировано 23 марта 2013 года.
- Helleday, Thomas. Pathways for mitotic homologous recombination in mammalian cells (англ.) // Mutation research : journal. — 2003. — November (vol. 532, no. 1—2). — P. 103—115. Архивировано 24 мая 2014 года.
- Luo, Guangbin; et al. Cancer predisposition caused by elevated mitotic recombination in Bloom mice (англ.) // Nature Genetics : journal. — 2000. — Vol. 26. — P. 424—429.
- Xu, T; GM Rubin. Analysis of genetic mosaics in developing and adult Drosophila tissues (англ.) // Development : journal. — 1993. — April (vol. 117, no. 4). — P. 1223—12237.
Литература
- Griffiths et al. 1999. Modern Genetic Analysis. W. H. Freeman and Company.
- Инге-Вечтомов С.Г. Генетика с основами селекции. — СПб.: Издательство Н-Л, 2010. — С. 193—194. — 718 с. — ISBN 978-5-94869-105-3.