Марита
Марита — половозрелая стадия гермафродитного поколения дигенетических сосальщиков (Digenea). В подавляющем большинстве случаев — паразит пищеварительной системы, полости тела или тканей позвоночных.
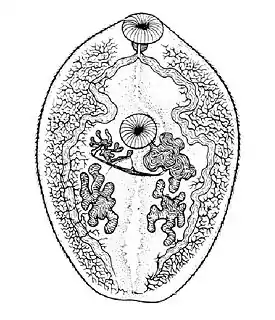
Строение
Размеры марит — особей гермафродитного поколения — варьируют от десятков микрометров до нескольких сантиметров. В отдельных случаях длина может достигать нескольких десятков сантиметров. Тело зачастую листовидной формы, уплощённое. Как правило, имеются две присоски: расположена на переднем конце тела ротовая, на дне которой находится ротовое отверстие, и брюшная (ацетабулюм), расположенная на брюшной стороне тела и обыкновенно смещена к переднему концу. В теле марит выделяют два отдела: локомоторный, или преацетабулярный (перед брюшной присоской); и генеративный, или постацетабулярный (после неё), которые несколько различаются по морфологии.
Однако в разных семействах существуют различные отклонения от такой общей схемы внешнего строения:
- Сем. Echinostomatidae вместо ротовой присоски имеется воротничок — складка на дорсальной стороне, частично заходящая на брюшную сторону.
- Сем. Paramphistomatidae: ротовая присоска полностью вторично редуцирована; терминально образуется передний орган (за счёт модификации самого переднего отдела пищеварительной системы). Брюшная присоска сильно смещена назад.
- Сем. Bivisiculidae: нет присосок вообще.
- Сем. Sanguinicolidae: нет брюшной присоски.
- Сем. Notocotylidae: также нет ацетабулюма, но развиваются вторичные прикрепительные образования (группы секреторных клеток).
- У высших фуркоцеркных трематод ротовая присоска очень маленькая, брюшная — почти редуцитрована; функцию прикрепления берёт на себя орган Брандеса.
- Сем. Bucephalidae: присоски отсутствуют; ротовое отверстие — на вентральной стороне, приблизительно посередине; терминально спереди — передний орган.
Кожно-мускульный мешок
Покровы трематод представлены типичным тегументом: сплошная наружная синцитиальная пластинка связана цитоплазматическими мостиками с погружёнными ядросодержащими частями — цитонами. В цитоплазме синцитиальной пластинки зачастую имеются актиновые шипики. Поверхностные структуры (такие как микровилли и микротрихии) у большинства представителей отсутствуют.
Под синцитиальной пластинкой расположены ряды кольцевых, продольных и диагональных мышц. Необходимо отметить, что хорошо выраженные мышечные слои имеются только в локомоторном отделе тела мариты; в генеративном же мышечных волокон значительно меньше и расположены они неупорядоченно. Строение мышечной ткани у трематод достаточно архаично: мышечная клетка представлена миоцитоном и отходящими от него отростками, которые содержат сократительные волокна. Каждый миоцитон имеет от двух до десяти отростков.
Паренхима у трематод фактически отсутствует. То, что ранее называли клетками паренхимы, оказалось либо цитонами тегумента, либо миоцитонами.
Нервная система
Нервная система трематод является типичным примером ортогона. Имеются три пары продольных стволов (дорсальные, латеральные и вентральные), которые связаны поперечными комиссурами (связками). Из трёх пар стволов, как правило, наиболее развиты брюшные. На переднем конце имеется трапециевидный ганглий. Вперёд от него отходит некоторое количество нервных окончаний, иннервирующих мышцы ротовой присоски и органы чувств. От вентральных стволов к брюшной присоске также отходит система нервных стволиков, образующих в её районе сплетение. В генеративном отделе тела решётка ортогона утрачивает свою правильность, нервные стволы сильно утончаются, комиссуры между ними практически исчезают, количество нейронов резко падает.
Протонефридиальная система
У марит трематод в отличие от остальных плоских червей имеется мочевой пузырь с эпителизованными стенками и своей мускулатурой. Его отверстие расположено субтерминально на заднем конце тела и снабжено сфинктером. От мочевого пузыря к переднему концу тела идут два главных собирательных канала, по ходу часто образующие узелки — сильно извитые и плотно упакованные участки. Далее главные собирательные каналы делятся на передние и задние продольные собирательные каналы. От этих каналов идут множественные ответвления, от которых в свою очередь отходят от трёх до девяти тончайших капилляров, на концах которых расположены циртоциты — мерцательные клетки.
Основной функцией протонефридиальной системы, по-видимому, является осморегуляция. Выделительная же функция у трематод осуществляется аммонотелическим путём (то есть продуктом азотистого обмена является аммиак, который выделяется во внешнюю среду через покровы). Кроме того, у многих трематод формируется вторичная (резервная, лакунарная) выделительная система: на поздних этапах развития мариты возникают множественные анастомозы между правой и левой частями протонефридиальной системы, а также дополнительные выросты. Функции вторичной выделительной системы — распределительная (в её сосудах обнаруживается большое количество аминокислот, простых сахаров) и выделительная (на стадии инцистировавшейся метацеркарии она работает как почка накопления).
Дыхательная система
Специализированных органов дыхания нет. Паразиты используют анаэробный обмен.
Пищеварительная система
Пищеварительная система трематод замкнутая. Ротовое отверстие расположено на переднем конце тела на дне ротовой присоски (если таковая имеется). Оно ведёт в перфарингиальный отдел, после которого расположена мускулистая глотка. Далее идёт пищевод, который перед брюшной присоской разделяется на две слепозамкнутые ветви кишечника.
Кроме того, у многих крупных трематод (например, Fasciola hepatica) наблюдается дополнительное ветвление пищеварительной системы, поскольку она берёт на себя распределительную функцию.
Совершенно иначе устроена пищеварительная система у представителей семейства Bucepalidae:
- У них ротовое отверстие расположено на брюшной стороне тела. Оно ведёт в небольшую щелевидную полость, после которой идёт глотка. За глоткой следует мешковидная кишка.
- Таким образом семейство Bucepalidae относят к подклассу Gasterostomata и противопоставлют всем остальным трематодам (подкласс Prosostomata).
Половая система
Большинство марит трематод являются синхронными облигатными гермафродитами, то есть имеют и женскую, и мужскую половую систему. Однако среди трематод есть и раздельнополые: в семействах Schistosomatidae и Didymozoidae.
Женская часть половой системы
Как правило имеется один яичник (хотя он бывает вторично сегментирован). От него отходит яйцевод, в котором расположено множество различных клапанов, препятствующих прохождению обратно в яичник зрелых яйцеклеток, а также сперматозоидов. В стенке яйцевода имеется вырост — семяприемник (служит для хранения спермы). В яйцевод впадает проток желточника (желточник обычно представлен множественными фолликулами). Также от яйцевода отходит так называемый Лауреров канал, который открывается отверстием на брюшной или спинной стороне тела. Его функция состоит в выведении из яйцевода избытков желточных клеток и сперматозоидов.
После места впадения Лаурерова канала яйцевод резко расширяется и образует крупную овальную камеру — оотип. Оотип имеет сильно утолщённые стенки, к которым примыкают одноклеточные скорлуповые железы. Совокупность оотипа и скорлуповых желёз называют тельцем Мелиса.
Мужская часть половой системы
Обыкновенно в развитии закладываются два семенника, которые впоследствии могут фрагментироваться. Исключением являются представители семейства Monorchidae, имеющие один семенник.
От семенников отходят семявыносящие каналы, идущие вперёд до брюшной присоски и объединяющиеся там в семяпровод. На семяпроводе могут иметься два вздутия — семенные пузырьки. Дальше семяпровод впадает в сумку цирруса — копулятивного органа.
