Ионный транспортёр
Ионный транспортёр — трансмембранный белок, который перемещает ионы (или другие небольшие молекулы) через биологическую мембрану для выполнения множества различных биологических функций, включая клеточную связь, поддержание гомеостаза, производство энергии и т. д.[1] Существуют разные типы транспортёров, включая насосы, унипортёры, антипортёры и симпортёры. Активные транспортёры или ионные насосы — это переносчики, которые преобразуют энергию из различных источников, включая аденозинтрифосфат (АТФ), солнечный свет и другие окислительно-восстановительные реакции, в потенциальную энергию, двигая ион по градиенту его концентрации.[2] Эта потенциальная энергия может затем использоваться вторичными транспортёрами, включая транспортёры ионов и ионные каналы, для управления жизненно важными клеточными процессами, такими как синтез АТФ.
Эта статья посвящена в основном ионным транспортёрам, действующим как насосы, но транспортёры также могут перемещать молекулы посредством облегчённой диффузии. Облегчённая диффузия не требует АТФ и позволяет молекулам, которые не могут быстро диффундировать через мембрану (пассивная диффузия), диффундировать вниз по градиенту их концентрации через эти переносчики белка. Ионные транспортёры необходимы для правильного функционирования клетки, поэтому они изучаются исследователями с использованием различных методов. Ниже приведены некоторые примеры клеточной регуляции и методов исследования.[3]
Классификация
Суперсемейство ионных транспортёров включает в себя 12 семейств[4]. Это разделение по семействам являются частью системы транспортной классификации (ТК), которая используется Международным союзом биохимии и молекулярной биологии; белки сгруппированы по таким характеристикам, как транспортируемые субстраты, механизм переноса, используемый источник энергии и также путем сравнения последовательностей аминокислот, составляющих каждый белок. Наиболее важным объединяющим фактором является заряженная природа субстрата, которая указывает на перенос иона, а не нейтральных частиц. Переносчики ионов существенно отличаются от ионных каналов: каналы — это поры, которые проходят через мембрану, тогда как транспортёры — это белки, которые должны менять форму, чтобы открыться, из-за этого транспортёры перемещают молекулы намного медленнее, чем это делают каналы.
Электрохимический градиент, или градиент концентрации — это разница в концентрации химической молекулы или иона в двух отдельных областях. В состоянии равновесия концентрации ионов в обеих областях будут равны, поэтому, если есть разница в концентрации, ионы будут стремиться течь «вниз» по градиенту концентрации — то есть от высокой концентрации к низкой. Ионные каналы позволяют определённым ионам, которые войдут в канал, течь вниз по градиенту их концентрации, выравнивая концентрации по обе стороны от клеточной мембраны. Ионные каналы и переносчики ионов достигают этого за счет облегчённой диффузии, которая является одним из видов пассивного транспорта. Однако только ионные транспортёры могут также осуществлять активный транспорт, который включает перемещение ионов против градиента их концентрации, используя такие источники энергии, как АТФ. Затем эти ионы могут использоваться вторичными транспортёрами или другими белками в качестве источника энергии.[5]
Источники энергии
Первичные транспортёры
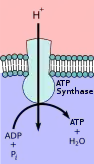
Первичные транспортёры используют энергию для переноса ионов, таких как Na+, K+ и Ca2+, через клеточную мембрану, и могут создавать градиенты концентрации. Этот транспорт может использовать АТФ в качестве источника энергии, или его можно использовать для генерации АТФ с помощью таких методов, как цепь переноса электронов в растениях.[5] This transport can use ATP as an energy source or it can be used to generate ATP through methods such as the electron transport chain in plants.[5]
Активные транспортёры
Транспортёры, использующие АТФ, преобразуют энергию АТФ в потенциальную энергию в виде градиента концентрации. Они используют АТФ для переноса иона из области низкой концентрации в область более высокой. Примерами белков, использующих АТФ, являются: АТФазы P-типа, которые переносят ионы Na+, K+ и Ca2+ путем фосфорилирования; АТФазы A-типа, переносящие анионы; ABC-транспортёры (переносчики кассет, связывающих АТФ), которые транспортируют широкий набор молекул. Примеры АТФазы P-типа включают Na+/K+-АТФазу, которая регулируется янус-киназой 2[6], Ca2+, АТФазу, а также Ca2+-АТФазу, которая проявляет чувствительность к концентрациям АДФ и АТФ. P-гликопротеин является примером ABC-транспорта, связывающего белки в организме человека.
АТФ-продуцирующие транспортёры
Транспортёры, продуцирующие АТФ, работают в противоположном направлении по отношению к транспортёрам, использующим АТФ. Эти белки переносят ионы от высокой к низкой концентрации, но в процессе образуется АТФ. Таким образом, потенциальная энергия в виде градиента концентрации используется для выработки АТФ. У животных этот синтез АТФ происходит в митохондриях с использованием АТФазы F-типа, иначе известной как АТФ-синтаза. Этот процесс использует цепь переноса электронов в процессе, называемом окислительным фосфорилированием. АТФаза V-типа выполняет функцию, противоположную АТФазе F-типа, и используется в растениях для гидролиза АТФ для создания протонного градиента. Примерами этого являются лизосомы, которые используют АТФазу V-типа для подкисления пузырьков или вакуолей растений во время процесса фотосинтеза в хлоропластах. Этот процесс можно регулировать с помощью различных методов, например, pH.[7]
Вторичные транспортёры

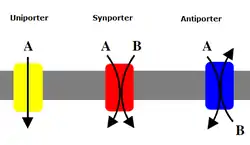
Вторичные транспортёры также переносят ионы (или небольшие молекулы) против градиента концентрации — от низкой концентрации к высокой, но, в отличие от первичных транспортёров, которые используют АТФ для создания градиента концентрации, они используют потенциальную энергию из градиента концентрации, создаваемого первичными транспортёрами, для транспортировки ионов. Например, натрийзависимый переносчик глюкозы, обнаруженный в тонком кишечнике и почках, использует градиент натрия, создаваемый в клетке натрий-калиевым насосом (как упоминалось выше), чтобы переносить глюкозу в клетку. Это происходит, когда натрий течет вниз по градиенту концентрации, обеспечивающему достаточно энергии, чтобы подтолкнуть глюкозу вверх по градиенту концентрации обратно в клетку. Это важно для тонкого кишечника и почек, чтобы предотвратить потерю глюкозы. Симпортёры, такие как натрий-глюкозный симпортёр, переносят ион с градиентом его концентрации, и они связывают транспорт второй молекулы в том же направлении. Антипортёры также используют градиент концентрации одной молекулы для перемещения другой вверх по градиенту концентрации, но связанная молекула транспортируется в противоположном направлении.[5]
Управление
Ионными транспортёрами можно управлять различными способами, такими как фосфорилирование, аллостерическое ингибирование или активация, и чувствительность к концентрации ионов. Использование протеина киназы для добавления фосфатной группы или фосфатаз для дефосфорилирования белка может изменить активность транспортёра. Будет ли белок активирован или ингибирован добавлением фосфатной группы, зависит от конкретного белка. При аллостерическом ингибировании регуляторный лиганд может связываться с регуляторным сайтом и либо ингибировать, либо активировать транспортёр. Ионные транспортёры также могут регулироваться концентрацией ионов (не обязательно тех, которые он переносит) в растворе. Например, цепь переноса электронов регулируется наличием ионов H+ (pH) в растворе.[5]
Методы изучения ионных транспортёров
Метод локальной фиксации потенциала
Метод локальной фиксации потенциала — это электрофизиологический метод, используемый для изучения каналов и переносчиков в клетках путем отслеживания протекающего через них тока. Этот метод был разработан Ходжкином и Хаксли до того, как стало известно о существовании каналов и транспортёров.[1][8]
Рентгеноструктурный анализ
Рентгеноструктурный анализ — удобный инструмент, который позволяет визуализировать структуру белков, однако это всего лишь снимок конформации одного белка. Структура транспортных белков позволяет исследователям лучше понять, как и что транспортёр делает для перемещения молекул через мембрану.[9]
Метод восстановления флуоресценции после обесцвечивания
Этот метод используется для отслеживания диффузии липидов или белков в мембране. Полезен для лучшего понимания подвижности транспортёров в клетке и её взаимодействия с липидными доменами и липидными рафтами в клеточной мембране.
Фёрстеровский резонансный перенос энергии
Метод, в котором флуоресценция используется для отслеживания расстояния между двумя белками. Используется для изучения взаимодействия транспортёров с другими клеточными белками[1]
Список транспортёров
| Ионные транспортёры |
|---|
| Транспортёр нейротрансмиттеров |
| Транспортёр глютаматов |
| Транспортёр моноаминов |
| ГАМК-транспортёры |
| Транспортёры глицина |
| Равновесные нуклеозидные транспортёры |
| Ca2+-АТФаза плазматической мембраны |
| Натрий-кальциевый обменник |
| Симпортёр хлорида натрия |
Примечания
- Maffeo C, Bhattacharya S, Yoo J, Wells D, Aksimentiev A (December 2012). “Modeling and simulation of ion channels”. Chemical Reviews. 112 (12): 6250—84. DOI:10.1021/cr3002609. PMC 3633640. PMID 23035940.
- Channels and Transporters // Neuroscience. — 2nd. — Sunderland, Mass. : Sinauer Associates, 2001. — ISBN 0-87893-742-0.
- Gadsby DC (May 2009). “Ion channels versus ion pumps: the principal difference, in principle”. Nature Reviews. Molecular Cell Biology. 10 (5): 344—52. DOI:10.1038/nrm2668. PMC 2742554. PMID 19339978.
- Prakash S, Cooper G, Singhi S, Saier MH (December 2003). “The ion transporter superfamily”. Biochimica et Biophysica Acta (BBA) - Biomembranes. 1618 (1): 79—92. DOI:10.1016/j.bbamem.2003.10.010. PMID 14643936.
- Fundamentals of biochemistry : life at the molecular level. — 2016-02-29. — ISBN 9781118918401.
- Hosseinzadeh Z, Luo D, Sopjani M, Bhavsar SK, Lang F (April 2014). “Down-regulation of the epithelial Na⁺ channel ENaC by Janus kinase 2”. The Journal of Membrane Biology. 247 (4): 331—8. DOI:10.1007/s00232-014-9636-1. PMID 24562791.
- Tikhonov AN (October 2013). “pH-dependent regulation of electron transport and ATP synthesis in chloroplasts”. Photosynthesis Research. 116 (2—3): 511—34. DOI:10.1007/s11120-013-9845-y. PMID 23695653.
- Swant J, Goodwin JS, North A, Ali AA, Gamble-George J, Chirwa S, Khoshbouei H (December 2011). “α-Synuclein stimulates a dopamine transporter-dependent chloride current and modulates the activity of the transporter”. The Journal of Biological Chemistry. 286 (51): 43933—43. DOI:10.1074/jbc.M111.241232. PMC 3243541. PMID 21990355.
- Shinoda T, Ogawa H, Cornelius F, Toyoshima C (May 2009). “Crystal structure of the sodium-potassium pump at 2.4 A resolution”. Nature. 459 (7245): 446—50. Bibcode:2009Natur.459..446S. DOI:10.1038/nature07939. PMID 19458722.