Гиббереллины
Гибберелли́ны — группа фитогормонов дитерпеновой природы, которые выполняют в растениях разнообразные функции, связанные с контролем удлинения гипокотиля, прорастания семян, зацветания и т. д. В контроле большинства морфогенетических процессов гиббереллины действуют в одном направлении с ауксинами и являются антагонистами цитокининов и абсцизовой кислоты (АБК).
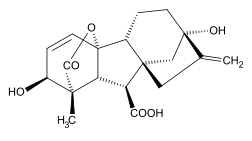
Химическая структура и классификация
Наиболее распространенными активными гиббереллинами являются GA1, GA3, GA4 и GA7 (от Gibberellic Acid), более слабой активностью обладают GA5 и GA6, которые тем не менее играют ключевую роль в гиббереллин-зависимой индукции цветения у однодольных. Среди прочих гиббереллинов большинство являются предшественниками в биосинтезе активных гиббереллинов либо продуктами их инактивации.
В отличие от ауксинов, критерием отнесения вещества к группе гиббереллинов является скорее соответствие определенной химической структуре, нежели наличие биологической активности. У растений, грибов и бактерий найдено 136 различных, близких по строению, веществ, относимых к группе гиббереллинов. Таким образом, гиббереллины — самый обширный класс фитогормонов. Гиббереллины представляют собой производные энт-гиббереллана и являются дитерпеноидами, однако предшественником биосинтеза служит энт-каурен. Гиббереллины могут иметь тетра- или пентациклическую структуру (дополнительное пятичленное лактонное кольцо) и соответственно содержат 20 (C20-гиббереллины, например ГК12) или 19 (C19-гиббереллины) атомов углерода. Большинство гиббереллинов — кислоты и поэтому принято обозначение ГК (гибберелловая кислота) с индексом, означающим порядок открытия, например ГК1, ГК3. Индекс никаким образом не отражает близость химической структуры или положения в метаболических путях. Несмотря на многообразие гиббереллинов, значительной биологической активностью обладает несколько соединений (ГК4, ГК1, ГК7, ГК3), остальные являются предшественниками биосинтеза или неактивными формами. В экспериментальной работе наиболее часто используется ГК3. Гиббереллины неустойчивы и быстро разрушаются в кислой или щелочной среде.
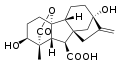
Гиббереллин А1 (ГК1) 
энт-Гиббереллан 
энт-Каурен
История открытия
Гиббереллины открыты японским учёным Е. Куросава (1926) при исследовании болезни риса (чрезмерном его росте), вызываемой грибом Gibberella fujikuroi Sow, поражающего посевы риса и вызывающего специфическое заболевание, при котором растения имеют аномально удлиненные междоузлия и низкую продукцию семян. В 1935 японский учёный Т. Ябута выделил гиббереллины из этого гриба в кристаллическом виде и дал им существующее название.
Содержание в растениях
У высших растений наиболее богаты гиббереллинами быстрорастущие ткани; они содержатся в незрелых семенах и плодах, проростках, развёртывающихся семядолях и листьях.
Биосинтез и инактивация гиббереллинов
Основное место синтеза гиббереллинов в растении — листовые примордии и молодые листья. Можно выделить три основных этапа биосинтеза гиббереллинов:
- Синтез энт-каурена из ГГДФ — катализируется ферментами группы терпен-синтаз (TPS): CPS (копалил-дифосфат-синтазой) и KS (энт-каурен-синтазой). Эти же ферменты принимают участие в синтезе фитоалексинов. Все ферменты TPS локализованы в пластидах.
- Синтез GA12 из энт-каурена — катализируется ферментами группы Р450-монооксигеназ: КО (энт-каурен-оксидазой) и КАО (оксидазой энт-кауреновой кислоты). Этот этап биосинтеза гиббереллинов проходит в эндоплазматической сети.
- Синтез прочих гиббереллинов из GA12 происходит в цитозоле и катализируется ферментами группы 2-оксоглютарат-зависимых диоксигеназ (2ODD). Среди ферментов 2ODD следует выделить GA-20-оксидазы и GA-3-оксидазы, которые осуществляют синтез биологически активных GA1, GA3, GA4 и GA7; а также GA-2-оксидазы, катализирующие реакции инактивации гиббереллинов (например, превращение активного GA1 в неактивный GA8).
У арабидопсиса и риса каждый из ферментов, катализирующих первые этапы биосинтеза гиббереллинов, кодируется одним-двумя генами: например, в геноме арабидопсиса имеется по одному гену, кодирующему ферменты CPS, KS и KO. Потеря функции этих генов у мутантов ga1, ga2 и ga3 соответственно приводит к серьезному сокращению уровня гиббереллинов и развитию фенотипа, типичного для гиббереллин-дефицитных мутантов — это карлики с низкой плодовитостью, без добавления экзогенных гиббереллинов их семена имеют низкую всхожесть, а взрослые растения не формируют цветоноса. В то же время ферменты 2ODD, действующие на последнем этапе биосинтеза гиббереллинов, кодируются большими мультигенными семействами, члены которых обладают ткане- и органоспецифическим характером экспрессии, а также по-разному регулируются в зависимости от внешних условий и стадии онтогенеза. Мутации с потерей функции любого из этих генов имеют более слабое фенотипическое проявление.
Основными путями инактивации гиббереллинов являются 2β-гидроксилирование с помощью GA-2-оксидаз; эпоксидирование с помощью фермента из группы цитохром-Р450-монооксигеназ EUI (Elongated Uppermost Internode, назван по фенотипу мутанта с потерей функции соответствующего гена) и метилирование с использованием S-аденозил-метионина как донора метильных групп — эту реакцию катализируют ферменты GAMT1 и GAMT2 (GA Methyl Transferase). По-видимому, все эти пути инактивации имеют одинаково важное значение в регуляции пула активных гиббереллинов в растении, так как потеря функции любого из перечисленных генов у мутантов eui, gamt1 и gamt2, а также у трансгенных растений с косупрессией генов GA2ox ведет к повышению концентрации активных гиббереллинов в десятки раз. Кроме того, существует слабо изученный путь инактивации гиббереллинов за счет образования конъюгатов с глюкозой — GA-глюкозильных эфиров.
Пути регуляции уровня активных гиббереллинов в тканях растений
Последний этап биосинтеза — синтез активных гиббереллинов из GA12 — играет ключевую роль в контроле уровня активных гиббереллинов в растении. Действительно, повышение уровня экспрессии гена GA20ox у трансгенных растений арабидопсис вызывает резкое повышение уровня гиббереллинов в тканях и серьезные фенотипические изменения, тогда как сверхэкспрессия гена AtCPS приводит только к накоплению энт-каурена, энт-кауреновой кислоты и GA12. В связи с этим основным способом регулирования концентрации активных гиббереллинов в тканях растения является контроль экспрессии генов, кодирующих ферменты семейства 2ODD, разные представители которого катализируют как реакции активации гиббереллинов (синтез GA1, GA3, GA4 и GA7 из GA12, который катализируется GA-3 и GA-20-оксидазами), так и реакции их инактивации (2β-гидроксилирование GA1, GA3, GA4 и GA7, которое катализируется GA-2-оксидазами). В настоящее время изучены генетические механизмы регуляции уровня экспрессии генов 2ODD в разных органах растений, а также в зависимости от определенных факторов внешней среды.
Регуляция конечным продуктом. Важную роль в поддержании гомеостаза гиббереллинов играет регуляция их биосинтеза конечным продуктом. Например, у арабидопсис повышение концентрации активных гиббереллинов вызывает транскрипции генов AtGA20ox и AtGA3ox, а снижение их концентрации повышает уровень экспрессии этих же генов. С другой стороны, активные гиббереллины позитивно регулируют уровень экспрессии генов AtGA2ox, участвующих в их инактивации. Точные механизмы, лежащие в основе регуляции экспрессии вышеперечисленных генов в зависимости от концентрации активных гиббереллинов, неизвестны, но показано, что в этот процесс вовлечены компоненты пути передачи сигнала гиббереллинов: рецептор GID1, репрессоры гиббереллинового сигналинга DELLA-белки, F-бокс-содержащий белок SLY/GID2.
Регуляция другими гормонами. В контроле развития растений гиббереллины являются антагонистами цитокининов, этилена и АБК и действуют в одном направлении с ауксинами. Для некоторых групп фитогормонов установлена способность регулировать концентрацию активных гиббереллинов через контроль экспрессии генов 2ODD. Например, ауксины, которые выполняют сходные с гиббереллинами функции, могут регулировать концентрацию активных гиббереллинов путём позитивной регуляции уровня экспрессии генов GA3ox и негативной регуляции экспрессии генов GA2ox. Основную роль в ауксин-зависимом контроле концентрации активных гиббереллинов играет транскрипционный фактор NPH4/ARF7, который регулирует экспрессию генов 2ODD, напрямую связываясь с их промоторами. Антагонистом NPH4/ARF7 в этом процессе является транскрипционный репрессор MSG2/IAA19.
Регуляция концентрации гиббереллинов в разных органах растений. Помимо регуляции экспрессии генов 2ODD в зависимости от концентрации активных гиббереллинов и других гормонов, существуют также механизмы регуляции их экспрессии в разных органах растения; некоторые из них:
- В апикальной меристеме побега (ПАМ). Транскрипционные факторы KNOX напрямую репрессируют экспрессию генов GA20ox, контролирующих биосинтез гиббереллинов, и позивно регулируют экспрессию генов IPT, контролирующих биосинтез цитокининов. Гены KNOX экспрессируются в корпусе ПАМ, в результате баланс гиббереллинов и цитокининов в центральной части ПАМ смещен в сторону цитокининов, что приводит к активной пролиферации клеток без дифференцировки. На периферии ПАМ, в зоне закладки листовых примордиев, экспрессия генов KNOX подавляется, что приводит к экспрессии гена GA20ox, повышению концентрации гиббереллинов и дифференцировке клеток. В основании ПАМ экспрессируется ген GA2ox, контролирующий инактивацию гиббереллинов, что важно для поддержания конститутивно низкого уровня гиббереллинов в ПАМ.
- В семенах. В темноте, при отсутствии активного фитохрома, активируется белок PIL5, который негативно регулирует экспрессию гена GA3ox и позитивно — экспрессию гена GA2ox; снижение уровня активных гиббереллинов подавляет процесс прорастения. НА свету активация фитохромов вызывает инактивацию PIL5, усиление экспрессии GA3ox и подавление экспрессии GA2ox — в результате повышается уровень активных гиббереллинов, что приводит к прорастанию (регуляция светом).
Рецепция и передача сигнала гиббереллинов
Путь передачи сигнала при ответе на гиббереллины включает в себя четыре основных компонента:
- Рецептор GID1 — растворимый ядерный белок.
- Компоненты убиквитин-лигазного комплекса SCFSLY/GID2.
- Репрессоры ответа на гиббереллин — белки DELLA, относящиеся к семейству транскрипционных факторов GRAS.
- Гиббереллин-зависимые транскрипционные факторы GAMYB, которые относятся к MYB-семейству транскрипционных факторов.
При отсутствии гиббереллинов с промоторами гиббереллин-регулируемых генов связаны репрессоры транскрипции DELLA, глюкозаминтрансферазы SPY/SEC стабабилизируют DELLA-белки. При связывании молекулы гиббереллина с рецептором GID1 индуцируется его взаимодействие с DELLA-белком и F-бокс содержащим белком SLY/GID2. В результате DELLA-белок присоединяется к убиквитин-лигазному комплексу SCFSLY/GID2, что стимулирует его убиквитин-зависимый протеолиз. К промоторам гиббереллин-регулируемых генов присоединяется транскрипционный фактор GAMYB, который запускает их экспрессию.
Белки DELLA — репрессоры ответа на гиббереллин. Транскрипционные репрессоры семейства DELLA были первыми из обнаруженных компонентов пути передачи сигнала гиббереллинов. Установлено, что ответ растений на гиббереллины зависит от деградации DELLA-белков, которые при отсутствии сигнала конститутивно связаны с промоторами гиббереллин-регулируемых генов. Мутанты с потерей функции генов, кодирующих DELLA-белки, обладают повышенной чувствительностью к гиббереллинам, напротив, мутанты с усилением их функций и трансгенные растения со сверхэкспрессией генов DELLA-белков — карлики.
Белки DELLA — небольшое подсемейство белков, относящихся к семейству GRAS (от GA1, RGA, SCARECROW), к которому также принадлежат транскрипционные факторы SCARECROW и SHORT-ROOT, участвующие в поддержании стволовых клеток апикальной меристемы корня и формировании его радиальной структуры. Молекулы DELLA-белков включают в себя ДНК-связывающий домен GRAS на С-конце и уникальный для этого подсемейства домен DELLA на N-конце, необходимый для взаимодействия с рецептором GID1 и F-бокс-содержащим белком SLY1/GID2. Мутации по GRAS-домену приводят к потерям функции DELLA-белков и усиливают ответ на гиббереллин. В то же время делецин в DELLA-домене приводят к накоплению DELLA-белков и конститутивной репрессии передачи сигнала гиббереллинов.
Белок GID1 — рецептор гиббереллинов. По своей последовательности и структуре рецептор GID1 относится к семейству гормон-чувствительных липаз, но не имеет ферментативной активности. Активированный рецептор GID1 взаимодействует с DELLA-белками. В результате происходит изменение конформации DELLA-белка, что делает возможным присоединение к нему ключевого компонента убиквитин-лигазного комплекса SCF — F-бокс-содержащего белка SLY/GID2. Это взаимодействие стимулирует убиквитинирование DELLA-белков комплексом SCFSLY/GID2 с последующей их деградацией 26S-протеасомой — таким образом происходит депрессия ответа на гиббереллины.
Компоненты убиквитин-лигазного комплекса У SCF (Scp1-Cullin-F-box) включают в себя четыре белка с разными функциями, за присоединение к ним белков-мишеней отвечает F-бокс-содержащий белок.
Гиббереллин-зависимые транскрипционные факторы GAMYB. Относятся к MYB-семейству транскрипционных факторов, действуют в передаче сигнала гиббереллинов находятся ниже DELLA-белков и позитивно регулируют транскрипцию гиббереллин-зависимых генов. Первым из идентифицированных транскрипционных факторов этой группы был транскрипционный фактор GAMYB1 ячменя, действующий в алейроновом слое эндосперма и позитивно регулирующий гиббереллин-зависимую экспрессию генов α-амилаз. В дальнейшем было установлено, что у ячменя, арабидопсис и риса транскрипционные факторы группы GAMYB участвуют также в гиббереллин-зависимом контроле цветения и развития органов цветка. Например, у арабидопсис прямой мишенью одного из белков GAMYB — GAMYB33 — является ген идентичности органов цветка LEAFY; активация другого GAMYB-белка — GLABROUS1 — необходима для гиббереллин-зависимого развития трихомов.
Другие известные компоненты передачи сигнала гиббереллинов. К числу негативных регуляторов передачи сигнала гиббереллинов у арабидопсис относятся белки SPY (SPINDLY) и SEC (SECRET AGENT), которые принадлежат к семейству N-ацетилглюкозаминтрансфераз. Мутации с потерей функции соответствующих генов приводят к фенокопиям растений, обработанных гиббереллинами.
Предположительной функцией белков SPY и SEC является стабилизация белков DELLA путём их посттрансляционной модификации.
Гиббереллин-регулируемые гены
В геноме арабидопсис обнаружено более 500 гиббереллин-регулируемых генов, половина из которых регулируется гиббереллинами напрямую, через деградацию DELLA-белков и присоединение транскрипционных факторов GAMYB. К их числу относятся:
- Гены, участвующие в контроле различных онтогенетических процессов, — в частности, прорастания (гены α-амилаз) и цветения (ген LEAFY);
- Гены, контролирующие метаболизм гиббереллинов — GA20ox, GA3ox, GA2ox;
- Гены, контролирующие передачу сигнала гиббереллинов — GID1, гены убиквитин-конъюгирующих ферментов Е1 и убиквитин-лигаз Е3, гены транскрипционных факторов GAMYB;
- Гены, контролирующие метаболизм и передачу сигнала других фитогормонов — например, убиквитин-лигаза XERICO, играющая роль в метаболизме АБК.
Функции
К числу наиболее известных функций гиббереллинов относятся контроль прорастания семян, роста стебля в длину, перехода к цветению и развития органов цветка. В настоящее время изучены молекулярные механизмы реализации этих функций: все они связаны с деградацией DELLA-белков, опосредованной рецептором GID1.
Контроль прорастания семян
Созревание семян связано с накоплением питательных веществ в зародыше и эндосперме. При прорастании происходит лизис запасных макромолекул эндосперма различными гидролитическими ферментами. Основную функцию в гидролизе крахмала в эндосперме выполняют α- и β-амилазы: α-амилазы гидролизуют крахмал до олигосахаридов, которые затем превращаются в мальтозу с помощью β-амилаз[1].
Гены α-амилаз были первыми генами, для которых установлен прямой контроль экспрессии с помощью гиббереллин-зависимого транскрипционного фактора GAMYB. Таким образом, гиббереллины, выделяемые зародышем, вызывают запуск экспрессии генов α-амилаз в алейроновом слое, что приводит к лизису крахмальных гранул эндосперма и обеспечивает молодой проросток питательными веществами.
Обработка растений гиббереллинами стимулирует пролиферацию клеток междоузлий и их рост растяжением. Гиббереллин-зависимое повышение частоты митозов наиболее хорошо заметно в субапикальных районах побега при переходе к цветению у розеточных растений длинного дня, а также в интеркалярных меристемах риса, растущего в глубокой воде. При этом в апикальной меристеме побега гиббереллины выполняют строго противоположную функцию — ингибируют пролиферацию и стимулируют дифференцировку клеток, благодаря чему для нормального развития растения важно поддерживание конститутивно низкой концентрации гиббереллинов в ПАМ. Таким образом, гиббереллины могут выполнять противоположные функции в контроле развития разных меристем.
Регуляция зацветания
Согласно многочисленным исследованиям существуют четыре основных пути регуляции зацветания: путь длинного дня, отвечающий за переход к цветению при увеличении продолжительности светового периода; автономный путь, который контролирует зацветание как при длинном, так и при коротком дне; путь вернализации, запускающий цветение после кратковременного периода низких температур; и гиббереллин-зависимый путь, который наиболее важен для перехода к цветению в условиях короткого дня.
Все эти пути могут взаимодействовать между собой — например, у арабидопсис, который может цвести как при длинном, так и при коротком дне. В связи с ослаблением гиббереллин-зависимого пути контроля зацветания, гиббереллин-дефицитные ga-мутанты арабидопсис совсем не цветут при коротком дне, а также демонстрируют задержку цветения в условиях длинного дня.
В основе гиббереллин-зависимого контроля зацветания лежит позитивная регуляция экспрессии генов, играющих ключевую роль в интеграции процессов цветения: индуктора экспрессии гомеозисных генов цветка — гена LEAFY (LFY), основного инициатора цветения — гена FLOWERING LOCUS T (FT) и его мишени — гена SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1).
В промоторе гена LFY обнаружена GAMYB-связывающая последовательность, с которой напрямую взаимодействует транскрипционный фактор GAMYB33. Мутации по GAMYB-связывающей последовательности делают невозможным повышение уровня экспрессии LFY в условиях короткого дня. В то же время трансформация гиббереллин-дефицитных ga-мутантов геном LFY под конститутивным промотором запускает их цветение при коротком дне.
У растений длинного дня роль гиббереллинов в контроле зацветания менее понятна. У этих растений увеличение продолжительности светового периода стимулирует рост стебля в длину (особенно хорошо это заметно у розеточных растений) с последующей индукцией цветения. Причиной этого, по-видимому, является фитохром-зависимое повышение уровня экспрессии генов GA20ox, которое приводит к увеличению концентрации свободных гиббереллинов в побегах. На арабидопсис было показано, что повышение концентрации гиббереллинов в листьях приводит к усилению экспрессии гена FT, кодирующего белок-флориген, который синтезируется в листьях и в дальнейшем перемещается в ПАМ. К сожалению, молекулярные механизмы влияния гиббереллинов на экспрессию FT в настоящее время не выяснены.
Роль в регуляции перехода к цветению
- Фитохром-зависимое повышение концентрации GA20ox в листьях вызывает повышение концентрации гиббереллинов, которое, в свою очередь, вызывает повышение концентрации белка FT. Этот процесс может также регулироваться фотопериодом, через ТФ СО. Белок FT перемещается из листьев в ПАМ.
- Повышение уровня гиббереллинов в ПАМ вызывает усиление экспрессии гена SOC, который негативно регулируется DELLA-белками GAI и RGA. Этот процесс негативно регулируется ТФ KNOX, которые стимулируют экспрессию генов GA2ox в Rib-зоне ПАМ.
- Экспрессия гена LFY напрямую позитивно регулируется гиббереллин-зависимым ТФ GAMYB33. DELLA-белки GAI и RGA негативно регулируют этот процесс через miPHK 156.
- Белки SOC1 и AGAMOUS-LIKE 24 (AGL24) формируют гетеродимер и позитивно регулируют экспрессию гена LFY; ТФ LFY позитивно регулирует экспрессию генов SOC1 и AGL24.
Итак, функции гиббереллинов связаны со стимуляцией вегетативного развития растений (прорастание, рост стебля в длину) и генеративного развития (перехода к цветению). Гиббереллины действуют в одном направлении с ауксинами и стимулируют биосинтез и передачу сигнала друг друга; кроме того, системы их рецепции передачи сигнала организованы по одному плану (рецептора взаимодействуют с компонентами убиквитин-лигазных комплексов Е3 и индуцируют протеолиз транскрипционных репрессоров). Цитокинины и АБК, напротив, являются антагонистами гиббереллинов.
Получение и применение
Гиббереллины получают главным образом микробиологическим способом из продуктов жизнедеятельности грибов рода Fusarium.
Гиббереллины применяют в практике растениеводства для повышения выхода волокна конопли и льна, для увеличения размеров ягод у бессемянных сортов винограда, для повышения урожайности трав, стимуляции прорастания семян (обработка гиббереллинами нарушает состояние покоя тканей и оказывает стратифицирующее действие на семена; при естественном выходе семян из состояния покоя содержание эндогенных гиббереллинов повышается) и др. Так как гиббереллины вызывают резкое ускорение роста зелёной массы растений, применение их должно сопровождаться усилением питания растений.
Для ускоренного созревания томатов, черешни, яблок, а также для предотвращения вылегания злаковых культур, используют обработки растений веществами-ретардантами, тормозящими действие гиббереллинов, например, 2-хлорэтилфосфоновой кислотой (этефоном).
См. также
Примечания
- D. E. Bilderback. A Simple Method to Differentiate between α- and β-Amylase // Plant Physiology. — 1973-03. — Т. 51, вып. 3. — С. 594–595. — ISSN 0032-0889.
Ссылки
- В. В. Чуб, Рост и развитие растений
- plant-hormones.info
- A Companion to Plant Physiology, Fifth Edition by Lincoln Taiz and Eduardo Zeiger
- Безуглова О. С. Гиббереллины. Удобрения и стимуляторы роста. Дата обращения: 22 февраля 2015.