Вегетативная гибридизация
Вегетати́вная гибридиза́ция — постулируемое в рамках «мичуринской агробиологии» Т. Д. Лысенко и его последователей явление образования при прививках растений «вегетативных гибридов», сохраняющих при половом размножении модификации, приобретенные в результате прививок, и не отличающихся от истинных гибридов, получаемых скрещиванием. «Вегетативная гибридизация» служила для последователей Лысенко одним из центральных «доказательств» ложности классической генетики и хромосомной теории наследственности[1]:40-41.
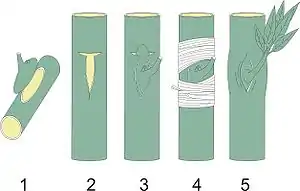
Понятие о вегетативных гибридах было введено И. В. Мичуриным, который использовал его вместо принятого к тому времени термина «прививочный гибрид». Первоначально вегетативной гибридизацией называли изменения привоя под влиянием подвоя при прививках растений.[2] Позже Лысенко и его последователи объединили под именем вегетативной гибридизации довольно разнородные явления, связанные, по их представлениям, с «расшатыванием» наследственности; они не только рекомендовали вегетативную гибридизацию как важный прием практической селекции, но и использовали данные по вегетативной гибридизации как аргумент, призванный опровергнуть хромосомную теорию наследственности и подтвердить их ламаркистские воззрения.[3]
В результате серии экспериментальных исследований 1930—1950-х годов наличие вегетативной гибридизации подтвердить не удалось, а развитие генетики во второй половине XX века показало ложность теоретических воззрений школы Лысенко. Поскольку образующийся в результате прививки или трансплантации организм относится не к истинным гибридам, а к химерам, в современной научной литературе термины «вегетативный гибрид» и «вегетативная гибридизация» практически не используются.
В учебнике Лобашева[4] термин «вегетативная гибридизация» рассматривается как эквивалент термина «прививочная гибридизация», а сам вопрос о влиянии подвоя на привой рассматривается в духе классической генетики. Более современные университетские учебники по генетике либо содержат жесткую критику теории вегетативной гибридизации, причём само название признается неудачной заменой более раннего термина «прививочный гибрид», которое тоже рекомендовано использовать лишь как иносказание,[5] либо вообще не упоминают ни о «прививочных», ни о «вегетативных» гибридах.[6] В монографии А. Е. Гайсиновича[7] по истории генетики понятие «вегетативной гибридизации» используется только при описании работ Лысенко и его сторонников.
С древнейших времён до XIX века

Использование прививок растений известно с античных времён и описано уже в работе Теофраста лат. De causis plantarum (О причинах [роста] растений). Теофраст утверждал, что подвой является лишь почвой для привоя и каждый из компонентов при срастании сохраняет свои особенности.
Вместе с тем, в некоторых случаях наблюдалось взаимное влияние привоя и подвоя. Степень влияния могла быть весьма различной, от незначительных модификаций (например, изменения плодовитости или периода вегетации) до образования побегов, в той или иной степени промежуточных по свойствам между подвоем и привоем.
Одним из первых случаев формирования побегов с промежуточными свойствами, получивших широкую известность, стал так называемый ракитник Адама. Когда французский садовник Адам в 1829 году привил пурпуровый ракитник Cytisus purpureus Scop. на желтоцветный ракитник Laburnum anagyroides Medik. (известный также под названиями Cytisus laburnum и Laburnum vulgare),[8] то на участке срастания возникли побеги с цветками промежуточной окраски с лепестками видоизмененной формы. Эти особенности сохранялись при вегетативном размножении. Полученный таким образом сорт ракитника и получил название Cytisus Adami (ракитник Адама). Вместе с тем, размножения семенами добиться не удалось, поскольку цветки ракитника Адама оказались практически бесплодны, а немногие полученные семена давали при посадке особи Laburnum anagyroides.[9]:262-264
Прививочные гибриды в работах Дарвина
В известном труде «Изменение животных и растений в домашнем состоянии» (англ. Charles Darwin «The Variation of Animals and Plants Under Domestication», 1868 год) Дарвин отвёл целый раздел этой проблеме, назвав его «Гибриды, происходящие вследствие прививки» (англ. Graft-hybrids).[10] Дарвин описал известные ему по данным литературы случаи образования помесей между отдельными видами или разновидностями без участия полового размножения. В существовании таких гибридов он был «убеждён» и считал такие факты «чрезвычайно важными» и способными «рано или поздно изменить взгляды физиологов на половое воспроизведение». (англ. „most important fact, which will sooner or later change the views held by physiologists with respect to sexual reproduction“)[10].

В большинстве рассматриваемых примеров Дарвин отметил неоднородность признаков тканей и органов прививочных гибридов с достаточно чёткими пространственными границами, склонность к возвращению частей со смешанными признаками к исходным родительским формам (реверсия). Однако, он считал, что «разделение подобного вида происходит отнюдь не редко … и у семенных гибридов» (англ. „But segregation of this kind occurs by no means rarely ... in seminal hybrids“).[10]
В качестве примеров прививочных гибридов Дарвин наряду с ракитником Адама рассматривал и Bizzarria (прививочный гибрид померанца с лимоном, известный с XVII века, для которого было характерно присутствие на одном растении как побегов и плодов исходных форм, так и «гибридных» плодов разнообразных форм: и со смешением признаков померанца и лимона, и состоявших из чётко различимых фрагментов, образованных тканями лимона и померанца), отметив, что Bizzarria размножается только черенками[11].
Дарвин рассмотрел различные гипотезы, объяснявшие возникновение и возможные причины вариабельности прививочных гибридов в рамках представлений о сохранении видовой (или сортовой) индивидуальности привоя и подвоя: изменение потока питательных веществ при прививке на подвой другого вида, возможное взаимное инфицирование привоя и подвоя, спонтанная изменчивости почек, ошибочно рассматриваемая как образование прививочного гибрида.
Подводя итог, Дарвин отметил, что во многих случаях — например, при прививках плодовых деревьев — наблюдаемое смешение признаков нельзя рассматривать как гибридизацию, но иногда — в частности, в опытах Магнуса с картофелем, случаи получения промежуточных форм при прививках «кажется, доказывают, что при некоторых неизвестных условиях гибридизация путём прививки возможна» (англ. Nevertheless, the cases above given seem to me to prove that under certain unknown conditions graft-hybridisation can be effected) и высказал предположение, что некие «элементы, которые идут на образование нового существа» могут содержаться не только в половых органах, но и в соматической ткани.[10]
Явления изменчивости при прививках Дарвин рассматривал лишь как один из штрихов общей картины разнообразия форм наследования и изменчивости одомашниваемых видов и проблемы соотношения передачи признаков при половом и вегетативном размножении. Результатом стало сделанное Дарвином в главе XXVII «Временная гипотеза пангенезиса» (англ. Provisional hypothesis of pangenesis) предположение, что во всех тканях организмов присутствуют субмикроскопические гранулы — геммулы, несущие наследственные признаки. Перенос геммул в репродуктивные органы, согласно этой гипотезе, — универсальный механизм, объясняющий весь комплекс наследственности и изменчивости. Дарвин предупреждал, что пангенез — не более, чем временная умозрительная гипотеза; вскоре после опубликования дарвиновской работы — в 1871 году в серии экспериментов по проверке существования геммул, поставленной Фрэнсисом Гальтоном (двоюродный брат Ч. Дарвина), гипотеза не подтвердилась. В этих опытах Ф. Гальтон переливал кровь от темноокрашенных кроликов светлоокрашенным, и никакого влияния на окраску шерсти у потомства не обнаружил[12].
Работы Винклера и Баура
Характер взаимного влияния подвоя и привоя долгое время оставался неясным. В начале XX века с развитием генетики началось тщательное изучение прививочных гибридов. Было показано, что в одних случаях речь идет о взаимном влиянии на уровне обмена метаболитами и гормонами,[9] в то время как в других — об образовании так называемых «химерных» побегов, состоящих отчасти из тканей подвоя, а отчасти — из тканей привоя. Механизмы образования промежуточных форм при прививке различных видов начал исследовать в 1907 году ботаник Ганс Винклер (впоследствии предложивший термин геном) в экспериментах по сращиванию томата Solanum lycopersicum и паслёна чёрного Solanum nigrum.[13]
В 1907 году Винклер предположил, что «клетки двух существенно различных видов могут сойтись другим, не половым путём, чтобы служить общей исходной точкой для организма, который при совершенно однородном общем росте одновременно будет обладать свойствами обоих первоначальных видов».
В 1912 году Винклер так классифицировал теоретические возможности скрещивания растений посредством прививки:
«Если задуматься над тем, как может путём прививки возникнуть гибрид, то, насколько я понимаю, представляются три теоретические возможности.
- Прежде всего было бы мыслимо, чтобы при прямом влиянии привоя на подвой, или наоборот, происходило бы такое изменение специфических признаков, которое привело бы к возникновению нового биотипа. Возникшие таким путём формы мы назовём прививочными гибридами влияния (Beeinflussungs-Propfbastarde) или модификационными прививочными гибридами (Modifications-Propfbastarde).
- Во-вторых, в месте срастания привоя и подвоя клетки, одни из которых происходят от привоя, другие — от подвоя, могут соединяться для образования адвентивного побега. Возникшие таким путём формы должны быть названы химерами.
- В-третьих, в месте срастания привоя и основного стебля может происходить более или менее похожее на процесс оплодотворения,
полное или частичное, слияние клеток привоя и подвоя, и продукт слияния может быть началом образования прививочного гибрида. Возникшие таким образом формы следует называть гибридами слияния или бурдонами».
На основе общетеоретических рассуждений (о неизменности зародышевой плазмы) Винклер отрицал возможность существования прививочных гибридов влияния. В 1912 году он писал: «до настоящего времени мы не знаем ни одного случая, доказывающего, или хотя бы делающего вероятной, возможность того, что при прививке один компонент сам по себе или в своём потомстве хоть сколько-нибудь изменяется в своих специфических свойствах под влиянием другого компонента. И нужно считать весьма вероятным, что такое прямое специфическое воздействие недостижимо путём прививки, Иными словами: модификационные прививочные гибриды, обусловленные взаимодействием компонентов, так же невозможны в действительности, как они невозможны по ходу наших рассуждений…»
Винклер направил свои усилия на то, чтобы показать, что настоящими прививочными гибридами могут быть только «гибриды слияния» (Burdonen). Методика опытов Винклера заключалась в том, что со стебля томата он удалял верхушку и все пазушные почки, и прививал на томат клином побег чёрного паслёна (либо наоборот — томат на паслён). Через 10—15 дней происходило полное срастание, и привой срезался на месте прививки с тем, чтобы декапитированное растение содержало небольшую часть привоя, зажатую с двух сторон подвоем. Поверхность среза образовывала каллюс, из которого вырастали придаточные почки, а из них — побеги. Значительная часть побегов представляла собой чистый паслен или томат, тогда как небольшая часть сочетала в себе четко различимые фрагменты паслена и томата. Такие побеги Винклер отчеренковывал, укоренял и выращивал до состояния взрослых растений.[1]:12
Для растений со смешанными признаками Винклер предложил термин «химеры», который с тех пор стал общепринятым.[9]:265-266 Винклер получил несколько химер различных видов, которые он подробно описал в своих работах (1907, 1910, 1912, 1916, 1938). Он выделил их в следующие формы:[1]:12
| Форма | Характеристика | Внешний вид |
|---|---|---|
| S. tubingense | Один слой клеток томата поверх клеток паслёна | По форме листьев близко к паслёну, с выраженным томатным опушением. |
| S. koelreuterianum | Один слой клеток паслёна поверх клеток томата | По форме листьев близко к томату, с поверхностью как у паслёна. |
| S. proteus | Два слоя клеток томата поверх клеток паслёна | Форма листьев паслёна, с сильно выраженными признаками томата |
| S. gaertnerianum | Два слоя клеток паслёна поверх клеток томата | Форма листьев томата, с сильно выраженными признаками паслёна |
| S. darwinianum | Смешанная форма (по мнению Винклера — бурдон) | Промежуточный между томатом и паслёном тип листьев; эпидермис сходен с эпидермисом паслёна. |
Против первоначального предположения Винклера о возможности гибридизации путём слияния вегетативных клеток выступил Эрвин Баур, изучавший пестролистные химеры. Он показал, что любые промежуточные формы образованы тканями исходных компонент, сохраняющих свою индивидуальность и морфологические признаки.[9]:265-271
Цитологический анализ позволил прояснить ситуацию и с другими известными случаями прививочных гибридов. В частности, выяснилось, что ракитник Адама представляет собой периклинальную химеру.[9]
Прививочные гибриды в работе селекционеров-практиков
Возможность взаимного влияния подвоя и привоя, приводящая к модификации свойств компонентов прививки, например, изменению периода вегетации, как и комбинирование привоя и подвоя с требуемыми свойствами, например, прививка европейских сортов винограда на устойчивый к «корневой тле» филлоксере подвой американских сортов, широко использовались в растениеводстве — в том числе и в селекционной работе для сохранения сортовых особенностей многолетних растений. Такие изменения при прививке происходят вследствие обмена метаболитами между привитыми компонентами и не наследуются при половом размножении, то есть являются проявлением модификационной изменчивости[14]
Американский селекционер Лютер Бёрбанк, выведший множество новых сортов растений[15][16], ставил эксперименты по изменчивости, получаемой в результате прививки. В своих опытах он, в частности, прививал пасленовые: при этом он наблюдал ряд модификаций (изменение периодов вегетации, формы и окраски плодов и листьев), он также отметил, что при прививке картофеля на томат клубни образовывались из ткани картофеля у границы тканей привоя и подвоя, но не на корнях томата-подвоя, так как, несмотря на прививку, ткани томата неспособны формировать клубни; эти клубни были мелки и имели измененную, по сравнению с нормальными клубнями форму. Бербанк — как и Баур — объяснял эти явления обменом метаболитов между привоем и подвоем, а изменения — «несоответствием» соков, которыми обмениваются привой и подвой с различающимися метаболизмами — «гибридизацией соков» (англ. sap hybridization)[17],[18].
С прививочными гибридами работал и французский биолог Люсьен Даниель, придерживавшийся взглядов о возможности «неполовой гибридизации» (фр. hybridation asexuelle) при прививках. Он экспериментировал с взаимными прививка различных культурных растений, среди которых была серия экспериментов по прививке топинамбура (Helianthus tuberosus) на подсолнечник (Helianthus annuus), проводившаяся им в течение 26 лет — с 1894 по 1921 г.; в течение этого периода Даниэль наблюдал лишь незначительные изменения привоя и подвоя. Лишь в 1921 г. ему удалось получить растение с клубневидными корни и «муфтой» у основания стебля, семена, полученные от привоя, дали стерильное, за исключением единственного растения, потомство. В свою очередь, среди потомства плодоносящего экземпляра, по сообщению Даниэля, оказались и формы с клубневидными корнями и «муфтой» у основания стебля, сохранявшие эти признаки до десятого поколения (последний посев в 1939 г.)[1]:36.
Возможность вегетативной гибридизации признавал и русский учёный-агробиолог М. В. Рытов (1846—1920). Он утверждал, что прививка — «сильное потрясение» для растения, которое, по его мнению, «равносильно и одинаково по своим результатам с тем, какое происходит при скрещивании от влияния пыльцы». Уклонения от прививки он считал хотя и «ничтожными», но при повторяющихся прививках — могущими усилиться и привести «даже к образованию нового сорта».[19]
«..Несмотря на все отрицательные мнения иностранных исследователей, не признающих влияния подвоя, я, на основании своих долголетних работ, буду категорически утверждать, что это влияние существует и при выводке новых сортов плодовых растений, с ним неизбежно приходится садоводу серьёзно считаться…»

Взаимовлияние компонентов прививки (метод ментора) использовал И. В. Мичурин при выведении новых сортов плодовых растений.
Мичурин считал некоторые свои сорта, в частности, гибрид яблони с грушей Ренет Бергамотный,[21] — вегетативными гибридами. Мичурин считал, что зарубежные критики вегетативной гибридизации — Фехтинг, Молиш, Кернер и др. «слишком односторонне и поверхностно» подошли к данному вопросу, имея, по его мнению, дело «со старыми, с установившейся наследственностью сортами, формами».[22]
Н. П. Дубинин в работе 1966 года высказал предположение, что Ренет Бергамотный — результат мутации.[23]
А. Жученко в выступлении на сессии Россельхозакадемии в 2005 году, посвященной 150-летию со дня рождения И. В. Мичурина, сказал: «Причём если в начале (до 1916 года) он (Мичурин) признавал возможность получения „вегетативных гибридов“, то в дальнейшем „отходит от такой односторонней и преувеличенной оценки роли подвоя…“».[24]
Один из крупнейших российских биологов XX века К. А. Тимирязев, ссылаясь на работы Дарвина, утверждал, что искусственное (при содействии человека) получение вегетативных гибридов, хотя и «подвергается сомнению» некоторыми исследователями, но все-таки «очень редко» встречается в случае прививок растений. В качестве примера таких фактов он привёл получение прививочного гибрида между боярышником и мушмулой, принадлежащими к разным родам. Тимирязев считал явления сложной наследственности (в некоторых случаях, прививки), однако, не такими полными, как при половом процессе, где, по его мнению, «происходит более тесное сочетание воспроизводящих элементов».[25]
Дискуссия о вегетативной гибридизации в СССР
Ряд русских селекционеров — в том числе и Мичурин — в начале XX века выражали сомнение в применимости генетики, в частности законов Менделя в практике селекционной работы, с достаточно резкой критикой теоретических положений Менделя и его последователей выступал и Тимирязев, противопоставивший «менделизм» дарвинизму.[26] В СССР, после прихода к власти Сталина, ситуация в науке вообще и биологии в частности оказались в значительной степени идеологизированной: в середине 30-х годов в рамках «классового подхода» к науке началась кампания по её идеологизации, при этом, опираясь на работу Сталина «Социализм или анархизм?» неодарвинизм Вейсмана, который был объявлен в этой работе уступающим неоламаркизму, был объявлен реакционным учением[27] — вместе с вейсмановской концепцией о передаче наследственных признаков только через зародышевую плазму половых клеток. В этой ситуации идея о наследовании приобретённых признаков прививочных гибридов становилась идеологически оправданной.
В конце 1930-х — 1940-е гг. вышел ряд работ Т. Д. Лысенко и его сторонников (А. А. Авакяна, И. Е. Глущенко и др.), популяризирующих идею вегетативной гибридизации.
«Менделисты-морганисты не могут со своих позиций допустить существования вегетативных гибридов. То, что никак нельзя было опровергнуть, относилось ими в разряд непонятных, необъяснимых явлений, названных химерами. На самом же деле, так называемые „химеры“ можно рассматривать как проявление смешанной наследственности, когда одна часть организма несёт свойства одного из компонентов, а другая — другого».
— Лысенко, 1946
Признанным лидером этого направления был И. Е. Глущенко, обобщивший учение о вегетативной гибридизации в монографии «Вегетативная гибридизация у растений» (1948).[28]
Суть лысенковской концепции вегетативной гибридизации состояла в том, что «природа» привоя изменяется в результате взаимодействия с подвоем в такой степени, что эти изменения начинают передаваться по наследству при размножении семенами. Вегетативная гибридизация рассматривалась Лысенко и его сторонниками как частный случай адекватного изменения организма в ответ на изменение окружающей среды и была связана с их представлениями об «ассимиляции» внешних условий (в данном случае, питательных веществ, поступающих из подвоя) живым организмом. Вегетативная гибридизация тесно увязывалась с лысенковской теорией «стадийного развития растений» — и выбор нужной стадии развития декларировался как необходимое условие получения «вегетативного гибрида».
Глущенко в своей монографии сформулировал основные посылки концепции следующим образом[1]:40-41:
Первое. …стадийно несфомировавшиеся организмы, непрошедшие ещё полного цикла развития, при прививке всегда будут изменять своё развитие, в сравнении с корнесобственными, то есть непривитыми растениями.
Второе. … Привой и подвой не могут обмениваться ни хромосомами ядер клеток, ни протоплазмой. И всё же наследственные свойства могут передаваться из подвоя в привой и обратно. Следовательно, пластические вещества, вырабатываемые привоем и подвоем, также обладают свойством породы, то есть наследственностью. Они обладают свойствами той породы, в которой они вырабатываются.
Третье. … Неправильным будет любое утверждение, говорящее о том, что свойство наследственности связано с каким-то особым веществом, в какой бы части организма или клетки оно не помещалось. Любая живая частичка или капелька тела … обладает свойством наследствености.
Четвёртое. Вегетативные гибриды принципиально не отличаются от гибридов, получаемых половым путём. Любой признак можно передавать из одной породы в другую посредством прививки так же, как и половым путём. …
В своей монографии Глущенко распространил понятие вегетативной гибридизации на эксперименты по трансплантации тканей у животных, пересадке ядер у одноклеточной водоросли ацетабулярии, трансформации у бактерий и даже на опыты зарубежных учёных (Нейгауз) по скармливанию личинкам дрозофилы среды, содержащей растёртые личинки соответствующего генотипа.[29] Во всех этих случаях предполагалось, что новые условия (пересаженная ткань, питательные вещества из тел личинок) будут «ассимилированы» организмом в целом и изменят его наследственность. Ключевым для признания возможности вегетативной гибридизации был вопрос о соотношении роли ядра и цитоплазмы в наследовании. Один из теоретиков «мичуринской генетики», Н. И. Фейгинсон подчеркивал:
… исключительное значение имеют результаты экспериментов по вегетативной гибридизации, которые показывают, что передача наследственных признаков и свойств может осуществляться через пластические вещества, без участия ядра.[30]
По Лысенко, вегетативная гибридизация была широко распространённым закономерным явлением:
… мы уже имеем все основания утверждать, что нет такой прививки стадийно молодого растения, которая не давала бы изменения наследственности.[31]
Вместе с тем, ряд генетиков и селекционеров с самого начала высказывались против концепции вегетативной гибридизации. В 1935—1939 гг. вышел ряд работ за авторством Н. И. Вавилова, Н. П. Дубинина, А. Р. Жебрака, А. И. Лусса, А. С. Серебровского, в которых концепция вегетативной гибридизации была подвергнута критике с теоретических позиций и эмпирической проверке. В ходе этих работ было показано, что вегетативная гибридизация в смысле, принятом в про-лысенковской литературе, не наблюдается, однако сторонники Лысенко отказались признать критику.[1]:9-10
Попытки дальнейшей критики работ по вегетативной гибридизации на растениях не имели большого успеха вплоть до середины 1960-х гг. Более того, авторы критических исследований сталкивались с трудностями при публикации своих результатов. Так, Ю. Я. Керкис, проверяя на томате открытые Менделем закономерности расщепления признаков при скрещивании, обнаружил, что в ряде случаев наблюдаются отклонения от теоретически ожидаемых расщеплений 3:1 и 1:2:1. Более тщательный анализ показал, что эти отклонения связаны с загрязнением исходного сортового материала томатов семенами, возникшими в результате случайного перекрестного опыления томатами других сортов. Эти результаты позволили ему поставить под сомнение чистоту методики проведения опытов по «вегетативной гибридизации» томатов, проводимых сторонниками Т. Д. Лысенко. Статья была включена в майский номер «Докладов АН СССР» за 1941 год, но была задержана и изъята из уже сверстанного журнала; опубликовать её удалось только в 1965 году после ликвидации монополии Лысенко в биологии.[32] По данным В. Я. Александрова, попытки экспериментального опровержения опытов по вегетативной гибридизации томатов привели к увольнению Ю. Я. Керкиса из Института генетики, в то время бывшего уже под началом Т. Д. Лысенко.[33]
С декабря 1952 года кампания по критике воззрений Т. Д. Лысенко и его последователей начала развиваться в «Ботаническом журнале», редколлегию которого, возглавляемую В. И. Сукачёвым, на тот момент контролировал Ботанический институт АН СССР. Дискуссия началась с вопросов видообразования, но позже охватила и другие стороны агробиологического учения Т. Д. Лысенко. В рамках этой кампании в 1955 году удалось опубликовать два обзора экспериментальных исследований вегетативной гибридизации, выполненных в ГДР. Ю. Л. Горощенко подготовил реферат работы немецкого селекционера Хельмута Бёме, а Д. В. Лебедев — статьи президента Академии сельскохозяйственных наук ГДР Ганса Штуббе.[34] Проведя несколько тысяч прививок на томатах и изучив несколько десятков тысяч потомков привитых растений, Х. Бёме и Г. Штуббе не получили ни одного подтверждения передачи признаков привоя при семенном размножении[35]. Основной вывод Штуббе гласил:
Исследования по проблеме вегетативной гибридизации растений, проведенные на большом материале в течение длительного времени, не дали никаких доказательств существования этого явления.[36]
Однако уже в 1958 году редколлегия журнала была расформирована и усилена сторонниками Лысенко, после чего критика вновь прекратилась.[37]
Критика приложения концепции вегетативной гибридизации к животным была несколько более успешна. В то время как ботаники вынуждены были прибегать к косвенной критике, перепечатывая работы своих зарубежных коллег, генетики животных изредка получали возможность публиковать результаты оригинальных исследований. Так, в 1957 году в работе Б. Л. Астаурова и В. П. Остряковой-Варшавер по андрогенным гибридам шелкопрядов, ещё раз подтвердившей тезис о решающей роли ядра в наследственности, были подвергнуты критике экспериментальные работы лысенковцев по трансплантации. Их статью удалось опубликовать только благодаря личной поддержке А. Е. Гайсиновича, в то время — члена редколлегии журнала «Известия Академии наук. Серия биологическая». Аналогичная критика была дана в монографии Н. Н. Соколова по межвидовым гибридам дрозофилы.[38]
С ликвидацией монополии Лысенко в биологии исследования по вегетативной гибридизации практически прекратились в виду полной бесперспективности её как метода «расшатывания наследственности» и «переделки природы растений». Точку в дискуссии поставила монография советского генетика С. Я. Краевого «Возможна ли вегетативная гибридизация растений посредством прививки?», увидевшая свет в 1967 году. Помимо обзора литературы, монография С. Я. Краевого содержала изложение результатов экспериментальных исследований, проделанных самим автором. В заключении книги С. Я. Краевой писал:
Большая и точная работа по прививкам с соблюдением строгого контроля как у нас, так и за рубежом показала, что прививки не являются фактором, вызывающим в сколько-нибудь значительных масштабах наследственную изменчивость растений. Кроме того, показана полная несостоятельность утверждения, что посредством прививок получаются вегетативные гибриды, аналогичные гибридам, получаемым от слияния половых клеток (гамет) в зиготу.[39]
Все, что осталось от этого обширного направления (по подсчётам Ж. Медведева, более 300 человек получили за работы по вегетативной гибридизации учёные степени, звания профессоров и академиков ВАСХНИЛ), на развитие которого ушло почти тридцать лет, — несколько форм томатов, для которых, к тому же, не было доказано возникновение в результате вегетативной гибридизации.[40] Это не отменяло работ с химерами и изучения взаимовлияния подвоя и привоя, но все они шли уже вне рамок учения о вегетативной гибридизации.
Взгляды современных последователей Лысенко и попытки реабилитации «вегетативной гибридизации»
В научной литературе концa XX — началa XXI века есть несколько статей, авторы которых интерпретируют свои экспериментальные данные с точки зрения наследуемых изменений при прививках растений[41][42][43][44][45]. Эти работы рассматривает профессор Ярослав Флегр[46] в публикации «Был ли Лысенко (частично) прав? Мичуринская биология в свете современной физиологии растений и генетики» (2002),[47], где он высказывает предположение, что, Лысенко и его последователи в отдельных случаях могли наблюдать — и абсолютно неверно интерпретировать — явления эпигенетического наследования, характеризуя сами воззрения Лысенко и его последователей следующим образом:
Мичуринская биология была собранием абсурдных теорий, как правило, основанных на анекдотичных наблюдениях или плохо спланированных опытах, проводимых без должного контроля и без какой-либо статистической обработки результатов.
Сторонником гипотезы возможности вегетативной гибридизации также является Юншэн Лю (Yongsheng Liu). В своей работе[48] он считает прививочную гибридизацию главным содержанием мичуринского учения. Упоминая критические работы в адрес этой теории[49], Liu пишет, что экспериментально было показано, что в некоторых случаях изменение характеристик при прививке стабильно и модификации, вызванные прививками, наследуются[50][51][52][53], что может быть вызвано горизонтальным переносом генов. При этом даже современные сторонники Лысенко, в частности, доктор Лю, признают: «Лысенко, там не менее, отвергал идею генов в качестве переносчиков информации», заменяя эту идею утверждением, что «эксперименты по вегетативной гибридизации предоставляют безошибочное доказательство того, что любая частица живого тела, даже соки, которыми обмениваются привой и подвой, обладают свойствами наследственности»[54].
Сторонниками взглядов Лысенко на вегетативную гибридизацию являются и ряд современных русскоязычных публицистов, которые пишут на тему истории советской науки в период правления И. В. Сталина[55][56][57].
Однако академическая наука в целом не разделяет эти взгляды. В частности, по факту появления статьи о вегетативной гибридизации в 8 томе Большой 62-томной энциклопедии «Терра»,[58] где эта концепция была упомянута как имеющая фактическое подтверждение, профессор кафедры экологии и защиты леса Московского государственного университета леса, доктор биологических наук С. С. Ижевский считает, что «текст статьи научно безграмотен, излагаемый в ней материал не подтверждается опытом селекционной науки».[59]
Химеры растений и прививочные гибриды

Согласно современным представлениям, взаимное влияние привоя и подвоя существует, однако является следствием объединения в физиологически единый организм генетически разнородных частей, то есть образование химеры различных типов. Химеры подробно обсуждаются в монографии Н. П. Кренке (1947). Современная трактовка содержится в более поздних публикациях[60].
При этом на формирование признаков компонентов химеры оказывает влияние транспорт метаболитов, выделяемых фрагментами (в том числе и фитогормонов)[61][62]. Однако поскольку генеративные органы химеры обычно формируются только подвоем, а в половом процессе участвуют гаметы, образующиеся только из одной из клеточных линий химеры, обмен генетическим материалом не происходит[14] и при размножении семенами признаки гибрида не наследуются. Типичным примером является ракитник Адама, семена которого дают исходный желтоцветный ракитник.
В случае пестролистных химер также может проявляться и пластидное наследование, описанное Бауром и Корренсом в начале XX века: так, в случае пестролистной ночной красавицы Mirabilis jalapa пластиды, синтезирующие хлорофилл, передаются с цитоплазмой яйцеклетки: так, при опылении цветков на белых (бесхлорофильных) побегах пыльцой цветков любых форм (белые, зелёные, пёстрые) из гибридных семян всегда вырастают белые, лишённые хлорофилла и вследствие этого нежизнеспособные растения; при опылении цветков на зелёных побегах картина противоположна (только зелёные); при опылении цветков на пестрых побегах гибридные семена дают все три формы[63].
Это явление служит подтверждением образования гамет из одной клеточной линии химеры и отсутствия внегенетической передачи признаков.
В современной литературе для химер, полученных прививкой, используется термин «прививочные гибриды». Взаимное влияние привоя и подвоя используется для усиления фенотипического проявления желаемых признаков, то есть модификаций, сохраняющихся при вегетативном размножении.
Ограниченная передача генов пластид между привоем и подвоем
В 2009 году в журнале Science была опубликована статья Ральфа Бока (англ. Ralph Bock) и Сандры Стегеманн (англ. Sandra Stegemann), в которой доказана возможность обмена генами между привоем и подвоем двух трансгенных линий табака,[64][65] несущих хлоропластные и ядерные маркеры. Верхушку растения одной из линий использовали в качестве привоя. После срастания привоя и подвоя участок растения, прилежащий к зоне срастания, вырезали и получали из него культуру клеток. В 45 случаях из 74 из вырезанного участка ткани были получены клеточные линии, содержащие оба маркера (ядерный маркер одного из сортов и хлоропластный маркер другого сорта). Авторы доказали, что происходила передача генов хлоропластов (либо больших участков хлоропластного генома, либо целых пластид) в обоих направлениях (от привоя к подвою и наоборот). Передачи ядерных генов не наблюдали ни в одном из случаев. Показано также, что это явление не связано со слиянием клеток привоя и подвоя и с разрезанием тканей растений. Из клеточных линий, содержащих оба маркера, были выращены взрослые растения. Из их семян выросли растения с тем же признаком (двойной устойчивостью к антибиотикам), что и у «гибридных» клеток, образовавшихся на границе привоя и подвоя. Таким образом, новая комбинация признаков, полученная в результате «вегетативной гибридизации», действительно может стать наследуемой, по крайней мере в лабораторных условиях.
Однако при этом авторы статьи отмечают, что хотя эти «данные демонстрируют обмен генетическим материалом между прививаемыми растениями, они не поддерживают догмат лысенкоизма о том, что „прививочная“ гибридизация была бы аналогичной гибридизации путём полового процесса»[64]. Вместо этого «открытие того, что перенос генов ограничен зоной контакта между привоем и подвоем, указывает на то, что данные изменения могут наследоваться только путём формирования бокового побега из зоны прививки».[64] Все другие побеги будут содержать только гены одного из исходных растений. Более того, неизвестно, могут ли подобные процессы происходить в природе. «Ясно лишь, что это должно происходить гораздо реже, чем предполагали сторонники лысенкоизма».[65] Кроме того, «вопреки взглядам лысенкоистов, „вегетативная гибридизация“ влияет только на крайне ограниченное число признаков, определяющихся геномами органелл — пластид и митохондрий», но не ядерными генами.[65]
Молекулярный биолог Николь Раск (англ. Nicole Rusk) так пишет о работе Ральфа Бока и Сандры Стегеманн: «Демонстрируя данные по переносу генов при прививках лишь на ограниченное расстояние, Бок представляет также молекулярные данные о том, что концепция вегетативной гибридизации является нежизнеспособной… Сегодня догмат Лысенко в значительной степени дискредитирован, хотя существует несколько опубликованных сообщений, заявляющих о наблюдениях наследуемых изменений в растении после процедуры прививки. Согласно Боку, эти сообщения поступают, в основном, от растениеводов, но молекулярные данные в пользу вегетативной гибридизации в настоящее время отсутствуют».[66]
Примечания
- И. Е. Глущенко
- О введении термина И. В. Мичуриным см. Гершензон 1979: 243; термин «прививочный гибрид» (калька с нем. Propfbastard и англ. graft-hybrid) использовался, например, в Баур, 1913.
- О распространении концепции вегетативной гибридизации на пересадку органов у животных и трансформацию у бактерий см. Глущенко, 1948: с. 29—72
О теоретическом значении вегетативной гибридизации см. Глущенко, 1948: с. 7.
О лидирующей роли Глущенко в вопросе о вегетативной гибридизации см. Александров, 1992: с. 158—160. - Лобашев, 1967: глава «Фенотипические изменения у прививочных гибридов и пересадка тканей», с. 584—589
- Гершензон, 1979: глава «Модификации и норма реакции», в особенности с. 243—245
- Дубинин, 1986; Айала, Кайгер, 1988
- Гайсинович, 1988: с. 268, 270, 334—335
- Синонимику Laburnum anagyroides Medik. см. в базе данных Germplasm Resources Information Network. Архивная копия от 26 октября 2012 на Wayback Machine
- Баур, 1913
- Charles Darwin «The variation of animals and plants under domestication»: Chapter XI
- Дарвин, Чарльз. Изменение животных и растений в домашнем состоянии = The Variation of Animals and Plants Under Domestication. — М.—Л.: ОГИЗ-Сельхозгиз, 1941. — С. 275.
- Пангенезис // Большая советская энциклопедия : [в 30 т.] / гл. ред. А. М. Прохоров. — 3-е изд. — М. : Советская энциклопедия, 1969—1978. (Дата обращения: 15 сентября 2011)
- П. А. Кошель «Вегетативное размножение растений» Электронная версия газеты «Биология» Архивная копия от 11 января 2012 на Wayback Machine
- Внучкова В. А. Вегетативная гибридизация // БСЭ (Интернет-публикация проверена 6 апреля 2008).
- Luther Burbank’s Plant Contributions by Walter L. Howard, University of California, Bulletin 691, March 1945
- «Intellectual Property Law for Engineers and Scientists», Howard B. Rockman, 2004
- Luther Burbank. Partner of Nature (Chapt. XVIII «In the Vegetable Garden»), NY, London: 1939 Архивная копия от 25 февраля 2009 на Wayback Machine (недоступная ссылка с 11-05-2013 [3209 дней])
- Luther Burbank. The Tomato and an Interesting Experiment: A Plant which Bore Potatoes Below and Tomato Above
- Рытов М. В. «Плодовый питомник». Кооперативное издательство. Москва—Ленинград, 1925 г., цит. по И. Е. Глущенко «Вегетативная Гибридизация», 1948 г., с. 34. Архивированная копия (недоступная ссылка). Дата обращения: 9 сентября 2010. Архивировано 9 сентября 2010 года..
- И.В.Мичурин «Применение менторов при воспитании сеянцев», 1916
- И. В. Мичурин «Итоги шестидесятилетних работ», Часть 1. «Принципы и методы работы», М: ОГИЗ Сельхозгиз, 1949 г.
- И. В. Мичурин «Ошибочное суждение многих учёных исследователей по признанию возможности явления вегетативных гибридов». Цит. по Глущенко, с. 32 (см. список литературы)
- Н. П. Дубинин Теоретические основы и методы работ И. В. Мичурина. М.: Просвещение, 1966. 183 с.
- Жученко А. А. Роль научного наследия И. В. Мичурина в повышении эффективности отечественного садоводства // Научная сессия Россельхозакадемии, посвященная 150-летию со дня рождения И. В. Мичурина (Интернет-публикация проверена 6 апреля 2008).
- К. А. Тимирязев «Исторический метод в биологии», Сочинения, т. VI, Сельхозгиз, 1939. Цит. по И. Е. Глущенко «Вегетативная Гибридизация» Архивированная копия (недоступная ссылка). Дата обращения: 9 сентября 2010. Архивировано 9 сентября 2010 года., с.30.
- Тимирязев К. А. Мендель, Грегор // Энциклопедический словарь братьев Гранат, 7-е издание, том 28, стр. 443—455, Изд-во бр. Гранат и Ко, СПБ, 1913
- Презент И. И. «Программа по курсу „Эволюционного учения“ для биологов 1-го курса ЛГУ» Архивная копия от 31 марта 2010 на Wayback Machine // Хрестоматия по эволюционному учению, Ленинград, 1934 (Интернет-публикация проверена 6 апреля 2008).
- Глущенко, 1948: с 29—72. О лидирующей роли Глущенко в вопросе о вегетативной гибридизации см. Александров, 1992: с. 158—160.
- О трактовке экспериментов на животных и бактериях как «вегетативной гибридизации» см. Глущенко, 1948: с. 24-29.
Современную трактовку упоминавшихся Глущенко опытов Гриффитса, а также Эвери, Мак-Леода и Мак-Карти по трансформации бактерий, см., например, в Гайсинович, 1988: с. 344. - Фейгинсон, 1963: с. 29 (цит. по Александров, 1992: с. 158).
- Лысенко Т. Д. Заключительное слово на 10-м заседании сессии ВАСХНИЛ 7 августа 1948 г.
- Гайсинович, 1988: с. 268.
- Александров, 1992: с. 159.
- См. Горощенко, 1955; Лебедев, 1955.
- Hagemann R (2002) «How did East German genetics avoid Lysenkoism?», Trends in Genetics 18, 320—324
- Лебедев 1955: с. 604.
- Об истории с публикацией данных Г. Штуббе см. Александров, 1992: с. 159—160.
О роли «Ботанического журнала» в борьбе с лысенковщиной в середине 1950-х см. также Медведев, 1993: 192—197 (главы «Возобновление биолого-агрономической дискуссии в 1952—1958 годах. Моральная победа „Ботанического журнала“» и «Разгром редакции „Ботанического журнала“ в декабре 1958 года и новый запрет на критику Т. Д. Лысенко»). - Гайсинович, 1988: с. 334—335. Имеются в виду работы Астауров Б. Л., Острякова-Варшавер В. П. Получение полного гетероспермного андрогенеза у межвидовых гибридов шелковичного червя: Экспериментальный анализ соотносительной роли ядра и цитоплазмы в развитии наследственности // Изв. АН СССР. Сер. биол. 1957. С. 157—175. Соколов Н. Н. Взаимодействие ядра и цитоплазмы при отдаленной гибридизации животных. М.: Изд-во АН СССР, 1959. 147 с.
- Краевой, 1967: с. 137.
- Оценку эффективности см. Медведев, 1993: с. 270—271.
- TAG Theoretical and Applied Genetics (недоступная ссылка) Taller J., Hirata Y., Yagishita N., Kita M. and Ogata S. 1998. Graft-induced Changes and the Inheritance of Several Characteristics in Pepper (Capsicum annuum L.). Theor. Appl. Genet. 97:705-713
- Fan, S.-Y. 1999. Phenotype Variation by the Action of Scion Prunus japonica. Thunb on Stock Prunus armeniaca L. Hereditas (Beijing) 21:43-44
- Hirata Y.: Graft-induced changes in eggplant (Solanum melongena L.) I. Changes of the hypocotyl color in the grafted scions and in the progenies from the grafted scions. Japan.J.Breed., 1979, 29:318-323.
- Hirata Y.: Graft-induced changes in skin and flesh color in tomato (Lycopersicon esculentum Mill.). J.Japan.Soc.Hort.Sci., 1980, 49:211-216.
- Hirata Y: Graft-induced changes in eggplant (Solanum melongena L.). II. Changes of fruit color and fruit shape in the grafted scions and in the progenies from the grafted scions. Japan.J.Breed., 1980, 30:83-90.
- Prof. RNDr. Jaroslav Flegr, CSc.
- Jaroslav Flegr. Was Lysenko (Partly) Right? Michurinist Biology in the View of Modern Plant Physiology and Genetics. Rivista di Biologia / Biology Forum Vol. 95, No. 2. (g 2002), pp. 258—272.
- Yongsheng Liu «Lysenko’s Contributions to Biology and His Tragedies», 2004
- Hagemann, R. [2002], How Did German Genetics Avoid Lysenkoism. Trends in Genetics 18: 320—324.
- Shinoto, Y. [1955], Graft Experiments in Eggplant. Kagaku 25: 602—607.
- Frankel, R. [1956], Graft-induced Transmission to Progeny of Cytoplasmic Male Sterility in Petunia. Science 124: 684—685.
- Zu, D.-M. and J.-G. Li [1964], Vegetative Hybridization in Plants and Animals. Science publishing house, Beijing (in Chinese).
- Hirata, Y., S. Ogata, S. Kurita, G.T. Nozawa, J. Zhou and S. Wu [2003], Molecular Mechanism of Graft Transformation in Capsicum annuum L. Acta Hort. 625: 125—130.
- Liu Y., Li B. and Wang Q. (2009) «Science and politics», EMBO Reports 10, 938—939
- «Дело генетиков. Новая книга Сигизмунда Миронина». Архивная копия от 4 октября 2008 на Wayback Machine
- М. А. Кудрявцев «Политическая биология»
- М. В. Алексеева, «Морган И Лысенко (К итогам сессии ВАСХНИЛ 1948 Г.)»
- Большая энциклопедия: В 62 томах. — М.: ТЕРРА, 2006.— ISBN 5-273-00432-2. Статья «Вегетативная гибридизация»
- Алексей Куприянов "Мичуринские методы создания энциклопедий. Прививка второго издания БСЭ к «Большой энциклопедии „Терра“».
- Примеры публикаций о химерах: Лобашев, 1967: с. 584—589 и Гершензон, 1979: с. 242—245.
- Lang, A., Chailakhyan, M. K., and Frolova, I. A. 1977. Promotion and inhibition of flower formation in a dayneutral plant in grafts with a short-day plant and a long-day plant. Proc Natl Acad Sci U S A. 74(6): 2412—2416.
- Лабунская Е. А., Жигалова Т. В., Чуб В. В. Анатомическое строение листа пестролистного Ficus benjamina сорта Starlight и взаимное влияние фотосинтезирующего и акцепторного компонентов химеры. Онтогенез, том 38, № 6, Ноябрь-Декабрь 2007, С. 471—480
- R. Hagemann. A Short Survey of the History of Plastid Genetics (between 1909 and 1964) Endocytobiosis Cell Res. (2004) 15, 7-23.
- Stegemann S., Bock R. (2009) «Exchange of genetic material between cells in plant tissue grafts» Science 324, 649—651
- Элементы — новости науки: Один из постулатов лысенкоизма частично реабилитирован
- http://www.nature.com/nmeth/journal/v6/n7/full/nmeth0709-484.html Rusk N. (2009) «Grafting as a potent molecular tool», Nature Methods 6, 484
Литература
- Айала Ф., Кайгер Дж. Современная генетика. В 3 т. Пер. с англ. — М.: «Мир», 1987—1988. (Т. 1. — 1987. — 295 с.; т. 2. — 1988. — 368 с. ISBN 5-03-000495-5; т. 3. — 1988. — 335 с. ISBN 5-03-000496-3.)
- Александров В. Я. Трудные годы советской биологии: записки современника. — СПб.: «Наука», 1992. — 262 с. ISBN 5-02-025850-4.
- Баур Э. Введение в экспериментальное изучение наследственности. Пер. с нем. под ред. П. И. Мищенко с дополнениями и исправлениями, сделанными автором для предстоящего 2-го немецкого издания // Труды Бюро по прикладной ботанике. Год 6-й, приложение 8-е. — 1913. — VI + 342 с.
- Гайсинович А. Е. Зарождение и развитие генетики. — М.: «Наука», 1988. — 424 с. ISBN 5-02-005265-5.
- Гершензон С. М. Основы современной генетики. — К.: «Наукова думка», 1979. — 508 с.
- Глущенко И. Е. Вегетативная гибридизация растений. — М: ОГИЗ-Сельхозгиз, 1948. — 240 с.
- Горощенко Ю. Л. Реферат: Helmut Böhme. Untersuchungen zum Problem der genetischen Bedeutung von Propfungen zwischen genotypisch verschiedenen Pflanzen. Zeitschrift für Pflanzenzüchtung. B. 33, H. 4, 1954, S. 367—418 (Г. Бёме. К проблеме генетического значения прививок между генетически различными растениями) // Бот. журн. — 1955. — № 3. — С. 434—437.
- Дарвин Ч. Происхождение видов путём естественного отбора. — Л.: «Наука», 1991. — 539 с. ISBN 5-02-026708-2.
- Дарвин Ч. Изменение животных и растений в домашнем состоянии. — М.-Л.: ОГИЗ-Сельхозгиз, 1941. — 620 с.
- Дубинин Н. П. Общая генетика. Изд. 3-е, перераб. и доп. — М.: «Наука», 1986. — 560 с.
- Дубинин Н. П., Глембоцкий Я. Л. Генетика популяции и селекция. — М.: «Наука», 1967. — 591 с.
- Кренке Н. П. Химеры растений. — М.-Л.: Изд-во АН СССР, 1947. — 388 с.
- Краевой С. Я. Возможна ли вегетативная гибридизация растений посредством прививки? — М.: «Наука», 1967. — 143 с.
- Лебедев Д. В. Новые данные к вопросу о вегетативной гибридизации // Бот. журн. — 1955. — № 4. — С. 603—604.
- Лобашев М. Е. Генетика. Изд. 2-е, перераб. — Л.: Изд-во ЛГУ, 1967. — 751 с.
- Лысенко Т. Д. Заключительное слово на 10-м заседании сессии ВАСХНИЛ 7 августа 1948 г. // О положении в биологической науке. Стенографический отчет сессии Всесоюзной академии сельскохозяйственных наук имени В. И. Ленина 31 июля — 7 августа 1948 г. — М.: ОГИЗ-Сельхозгиз, 1948.
- Медведев Ж. А. Взлёт и падение Лысенко. История биологической дискуссии в СССР (1929—1966). — М.: «Книга», 1993. — 348 с. ISBN 5-7132-0005-1.
- Мичурин И. В. Итоги шестидесятилетних работ. 5-е изд. — М.: Сельхозгиз, 1949. — 672 с.
- Фейгинсон Н. И. Корпускулярная генетика. Критический обзор. — М.: Сельхозиздат, 1963. — 544 с.