Palaeocastor
Palaeocastor (лат.) — род ископаемых бобров, живших в Северной Америке в позднем олигоцене и раннем миоцене.
| † Palaeocastor | ||||||||
|---|---|---|---|---|---|---|---|---|
 | ||||||||
| Научная классификация | ||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Класс: Подкласс: Клада: Инфракласс: Надотряд: Грандотряд: Отряд: Подотряд: Инфраотряд: Надсемейство: Бобровые Семейство: Род: † Palaeocastor |
||||||||
| Международное научное название | ||||||||
| Palaeocastor Leidy, 1869 | ||||||||
| ||||||||
Внешний вид и палеоэкология
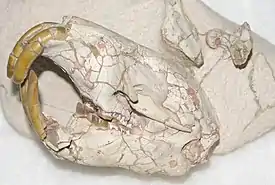
Представители рода Palaeocastor были грызунами среднего размера; их рост составлял около 12, а суммарная длина тела — около 30 см[1]. Масса тела этих грызунов составляла от 0,8 до 1,2 кг[2]. Лишь крупнейший из представителей рода, P. magnus, был, вероятно, крупней современных бамбуковых крыс, масса тела которых может достигать 4 кг[3]), а P. fossor (частью палеонтологов включаемый в род Fossorcastor) был размером с современную чернохвостую луговую собачку[4].

Строение тела представителей рода свидетельствует о приспособленности к рытью нор и жизни в них. Их череп был коротким, низким и широким, что характерно для других роющих грызунов (в частности, землекоповых). Резцы спереди уплощённые, нижняя челюсть в целом более примитивная, чем у современных бобров, со сдвинутой вперёд восходящей ветвью и не столь сильно, как у более современных видов, смещённым к средней оси нижнечелюстным мыщелоком. Более низкое расположение на черепе подглазничного отверстия также является примитивным по сравнению с современными бобрами и даже с менее древними представителями подсемейства Palaeocastorinae[5]. Надбровные дуги, сходясь, образуют умеренно развитый сагиттальный гребень в месте наименьшего расстояния между глазницами или выше него. Слуховые буллы (костные структуры, окружающие внешние выходы слухового канала) высокие и закруглённые[6]. Передние лапы мощные, широкие и уплощённые, хвост часто укороченный (для сравнения, у современных бобров уплощены и передние, и задние лапы, что является адаптацией к водному образу жизни, а хвост широкий и расплющенный, напоминающий по форме лопасть весла)[5].
Степень приспособленности к норному образу жизни у разных видов рода была разной — в частности, P. peninsulatus был менее специализирован, чем более поздний P. fossor, сохранив широкий зрительный канал, что свидетельствует о том, что он больше времени проводил на свету; кроме того, у разных видов наблюдаются черты, указывающие на разный основной способ рытья, и если P. fossor использовал для этого в основном зубы, то P. peninsulatus свои норы, по-видимому, рыл преимущественно лапами[7].

Род Palaeocastor обитал на территории Северной Америки в позднем олигоцене и раннем миоцене (30—20 млн лет назад). В это время в регионе, где найдены остатки представителей рода (современные штаты Небраска[8], Монтана и Орегон[7]), климат стал более прохладным и засушливым, господствующим типом местности стали семиаридные степи, что заставило многих грызунов адаптироваться к норному виду жизни. Своя норная ветвь, основу рациона которой составляли ненасыщенные кремнием травы, выделилась и среди бобровых[3]. По мнению Мартина, норные бобры были тупиковой боковой ветвью в своём семействе, и ни одна из последующих его ветвей от них не происходит[9].
Некоторые виды рода Palaeocastor рыли норы необычной спиральной формы, заканчивавшиеся отходящей под прямым углом спальной камерой (где обычно и находят их кости). В полностью сохранившихся норах количество витков может составлять от 6 до 12, а длина от 210 до 275 см, средний уклон от 25° до 30°, спальная камера горизонтальная или слегка наклонная, с самой низкой точкой у входа, вытянутая (сообщалось о камерах длиной до 4,5 м). Дополнительные выходы из спальной камеры отсутствуют, свидетельствуя, что хозяева нор предпочитали не спасаться бегством от хищников (которые из-за небольшого диаметра хода не могли быть намного крупней их самих), а занимать оборону у входа в жильё[4]. Необычная спиральная форма нор требовала бо́льших усилий, чем при рытье прямых наклонных ходов такой же длины. В качестве причин, приведших род Palaeocastor к такой форме, назывались территориальная скученность (представители рода жили большими колониями, подобно современным луговым собачкам[8], и прямые норы должны были часто пересекаться), лучшая защита от хищников, а также более надёжное сохранение температуры и влажности в спальной камере[10].
История открытия и классификация
Голотип рода был описан в 1856 году как Steneofiber nebrascensis; таким образом, этот американский ископаемый бобр был вначале причислен к роду своих евразиатских сородичей. В 1869 году Джозеф Лейди в работе, описывающей окаменелости из Южной Дакоты и Небраски, дал более подробное описание вида и определил его в качестве голотипа нового рода, которому дал имя Palaeocastor (буквально — «древний бобр»). Достоверно не известно, где именно были найдены остатки, по которым был описан голотип: это мог быть либо лептаухениевый костеносный слой на Уайт-Ривер, либо формация Арикари в окрестностях Розбада в Южной Дакоте. В 1935 году Р. А. Стиртон, проводя ревизию рода Palaeocastor, назвал второй вариант более вероятным. Стиртон также уточнил датировку окаменелостей: если до этого их относили к олигоцену, то Стиртон датировал их ранним миоценом[6].

В 1891 году, когда геолог Эрвин Барбоур проводил экспедицию в бедленде в районе реки Уайт-Ривер (Небраска), местные жители обратили его внимание на необычные окаменелости. Они представляли собой наполненные песком спиральные трубки длиной до 3 метров со стенками из белого волокнистого материала. Такие окаменелости находили в местности, которая на тот момент считалась дном пересохшего древнего озера. Местное население называло их «чёртовыми штопорами» (англ. Devil's corkscrews), и Барбоур, полагая, что открыл новый, до этого неизвестный род ископаемых пресноводных губок, дал ему латинское название Dæmonelix, переведя название с английского[8].
В дальнейшем, когда стало ясно, что местность, где находили окаменелости Dæmonelix, была 22 миллиона лет назад не озером, а семиаридной степью, Барбоур предположил, что его находка была не губкой, а неизвестным наземным растением. Однако палеонтологи Эдвард Коп и Теодор Фукс, обратившие внимание на то, что в одном из «штопоров» найдены кости неизвестного грызуна, уже в 1893 году независимо друг от друга выдвинули гипотезу, согласно которой Daimonelix представляли собой не окаменелости организмов, а норы животных. В 1905 году Олаф Петерсон из Музея Карнеги подтвердил, что находки Барбоура были окаменелыми норами, а грызуны, их рывшие — ископаемыми родичами бобров. Белёсое вещество, из которого состояли стенки трубок, включало корни растений, благодаря высокому содержанию кремния в грунтовых водах превратившиеся в стеклянистую массу, сохранившую спиральную форму нор[8].
Петерсон описал найденные остатки бобров как два новых вида рода Steneofiber[11], но в дальнейшем они были отнесены к роду Palaeocastor. В последующие годы различными авторами выделялись многочисленные виды этого рода, в том числе P. fossor, P. nebrascensis, P. barbouri, P. magnum, P. milleri, P. peninsulatus и P. simplicidens[6][8][12][13], часть из которых была в итоге выделена в отдельные таксоны (Pseudopalaeocastor, Fossorcastor), а другие определены как синонимичные. В 1987 году Ларри Мартин выделил в семействе бобровых новое подсемейство Palaeocastorinae, к которому отнёс четыре известных рода ископаемых бобров (Palaeocastor, Capatanka, Capacikala и Euhapsis) и два новых. Рода Palaeocastor и Capatanka в классификации Мартина образуют трибу Palaeocastorini. В качестве общих признаков для этого подсемейства, представители которого были адаптированы для рытья нор и жизни в них, Мартин выделил уплощённые резцы и короткий, круглый в сечении хвост[14]. Видовая и даже родовая классификация внутри подсемейства Palaeocastorinae с тех пор оставалась проблематичной, и в 2010 году Клара Стефен указывала, что эту классификацию трудно убедительно обосновать такими характеристиками, как форма зубов и форма и размер черепа[15].
Примечания
- Agate Fossil Beds: Agate Fossil Beds National Monument, Nebraska. — Interior Department, National Park Service, Division of Publications, 1989. — P. 69. — (National Park Handbook Series). — ISBN 9780912627045.
- Dietland Muller-Schwarze. The Beaver: Its Life and Impact. — 2nd Ed. — Ithaca, NY: Cornell University Press, 2011. — P. 8. — ISBN 978-0-8014-5010-5.
- J. X. Samuels, and B. Van Valkenburgh. Craniodental Adaptations for Digging in Extinct Burrowing Beavers // Journal of Vertebrate Paleontology. — 2009. — Vol. 29, № 1. — P. 254—268.
- L. D. Martin, and D. K. Bennett. The Burrows of the Miocene Beaver Palaeocastor, Western Nebraska, U.S.A. // Palaeogeography, Palaeoclimatology, Palacoecology. — 1977. — Vol. 22, № 3. — P. 173—193.
- William W. Korth. Comments on the Systematics and Classification of the Beavers (Rodentia, Castoridae) // Journal of Mammalian Evolution. — 2001. — Vol. 8, № 4. — P. 279—296.
- Macdonald, J. R. The Miocene faunas from the Wounded Knee area of western South Dakota // Bulletin of the American Museum of Natural History. — 1963. — Vol. 125. — P. 190—194.
- Jonathan Jean-Michel Calede. Skeletal Morphology of Palaeocastor peninsulatus (Rodentia, Castoridae) from the Fort Logan Formation of Montana (early Arikareean): Ontogenetic and Paleoecological Interpretations // Journal of Mammalian Evolution. — 2014. — Vol. 21, № 2. — P. 223—241. — doi:10.1007/s10914-013-9231-8.
- Martin, L. D. The Devil’s Corkscrew // Natural History. — 1994. — № 4. — P. 59—60.
- Korth, 1994, p. 145.
- Robert C. Meyer. Helical burrows as a palaeoclimate response: Daimonelix by Palaeocastor // Palaeogeography, Palaeoclimatology, Palaeoecology. — 1999. — Vol. 147, № 3—4. — P. 291—298.
- O. A. Peterson. Description of new rodents and discussion of the origin of Daemonelix // Memoirs of the Carnegie Museum. — 1905. — Vol. 2, № 4. — P. 139—202.
- Korth, 1994, p. 147.
- Samuels, 2007, p. 103.
- Korth, 1994, pp. 145, 147.
- C. Stefen. Morphometric considerations of the teeth of the palaeocastorine beavers Capacikala, Palaeocastor and "Capatanka" // Palaeontologia Electronica. — 2010. — Vol. 13, № 1. — P. 1—34.
Литература
- William W. Korth. The Tertiary Record of Rodents in North America. — New York: Springer Science+Business Media, 1994. — ISBN 978-1-4899-1444-6.
- Joshua Xavier Samuels. Paleoecology and Functional Morphology of Beavers (family Castoridae). — Los Angeles, CA: University of California, 2007. Архивная копия от 29 мая 2015 на Wayback Machine
Ссылки
- Castoridae: †Palaeocastorinae на сайте Mikko’s Phylogeny Archive (англ.)