Burianosaurus
Burianosaurus (лат.) — род травоядных динозавров из инфраотряда орнитоподов верхнемеловой эпохи Европы. Типовой и единственный вид Burianosaurus augustai назван и описан Мартином Мазухом и коллегами в 2017 году. Имя рода дано в честь чешского палеохудожника Зденека Буриана. Видовое имя дано в честь чешского палеонтолога и популяризатора науки Йозефа Аугусты[1]. Это были первые скелетные остатки динозавра в Чехии[2].
| † Burianosaurus | |||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Бедренная кость | |||||||||||||||||||||||||||||||||||||||||
| Научная классификация | |||||||||||||||||||||||||||||||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Надотряд: Отряд: Подотряд: † Цераподы Инфраотряд: Род: † Burianosaurus |
|||||||||||||||||||||||||||||||||||||||||
| Международное научное название | |||||||||||||||||||||||||||||||||||||||||
| Burianosaurus Madzia et al., 2017 | |||||||||||||||||||||||||||||||||||||||||
| Единственный вид | |||||||||||||||||||||||||||||||||||||||||
|
† Burianosaurus augustai Madzia et al., 2017 |
|||||||||||||||||||||||||||||||||||||||||
| Геохронология Сеноманский век 100,5—93,9 млн лет
◄ Наше время
◄ Мел-палеогеновое вымирание ◄ Триасовое вымирание
◄ Массовое пермское вымирание
◄ Девонское вымирание
◄ Ордовикско-силурийское вымирание
◄ Кембрийский взрыв
|
|||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
История исследования
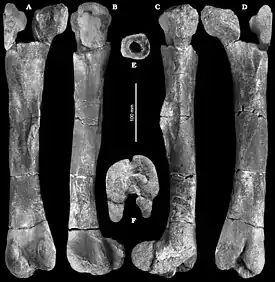
Голотип NMP Ob 203, представляющий собой хорошо сохранившуюся почти целую левую бедренную кость, обнаружен в слоях формации Peruc-Korycany, датированных поздним сеноманом, недалеко от города Кутна-Гора, Среднечешский край, Чехия[1].
Голотип был обнаружен Мартином Мазухом в 2003 году[2].
Описание

Burianosaurus — это орнитопод, не входящий в кладу Ankylopollexia и отличающийся от других орнитопод, не входящих в эту кладу, по следующим сочетаниям уникальных признаков: грацильная бедренная кость (возможно, как у Eousdryosaurus, но отличается от всех неигуанодонтовых орнитопод, Rhabdodontomorpha, тенонтозавра, Elrhazosaurus и Valdosaurus), тело кости прямое в медиальной и латеральной проекциях (как у Zalmoxes robustus и Zalmoxes shqiperorum, но отличается от неигуанодонтовых орнитопод, Mochlodon, Muttaburrasaurus, Rhabdodon sp., тенонтозавра, Eousdryosaurus, Elrhazosaurus и Valdosaurus), место прикрепления длинной хвостовобедренной мышцы (m. caudofemoralis longus) находится у основания четвёртого вертела (отличается от всех дриозаврид), в медиальной проекции дистальные мыщелки выступают над телом кости (как у всех неигуанодонтовых орнитопод, Mochlodon, Zalmoxes robustus, Eousdryosaurus, Elrhazosaurus, Valdosaurus и, возможно, Muttaburrasaurus и тенонтозавра, но отличается от Rhabdodon sp. и Zalmoxes shqiperorum), межмыщелковая борозда (для разгибателя) относительно глубокая и узкая (похоже на Elrhazosaurus и немного глубже, чем у Zalmoxes shqiperorum и, возможно, также Valdosaurus; но отличается от других Rhabdodontomorpha, неигуанодонтовых орнитопод, Dryosaurus и Eousdryosaurus).
Длина бедренной кости 400 мм[1].
Палеобиология
У экземпляра незрелая костная микроструктура во внутренней губчатой зоне без вторичных остеонов и зрелая пластинчатая зона с гаверсовой системой во внешнем слое. Таким образом, кость, вероятно, принадлежит взрослому животному[2]. На доступной в данное время фотографии (гистологический препарат отсутствует) видно, что наружный кортикальный слой состоит из неплотно упакованной гаверсовой системы, это предполагает относительную зрелость кости, вероятно, молодой взрослой особи. На имеющихся в распоряжении учёных изображениях отсутствуют линии замедления и остановки роста и внешняя фундаментальная система, но на этих изображения представлена только небольшая область кости. Подтверждение, что голотип не является ювенильной особью способствует включение его в филогенетический анализ, поскольку кодирование признаков не должно сильно зависеть от онтогенеза[1].
Филогения
Первоначально кость считалась принадлежащей неопределённому игуанодонтиду, маленький размер которого мог объясняться островной карликовостью[2].
Поскольку существуют различные гипотезы филогенетических взаимоотношений среди базальных Neornithischia, и, учитывая неполноту голотипа, для выяснения филогенетического положения Burianosaurus было использовано два набора данных. Первый основывался на исследовании Макдональда 2012 года[3] в модификации Boyd и Pagnac[4]. Целью включения Burianosaurus в этот анализ было выяснение, является ли динозавр членом клады Ankylopollexia. Итоговая матрица данных содержала 135 признаков для 64 таксонов. Полученное дерево плохо разрешено, потому что было кодировано только семь признаков Burianosaurus, а также большинство таксонов представлены фрагментарным материалом. Тем не менее, Burianosaurus находится за пределами Ankylopollexia внутри относительно хорошо разрешённой клады Rhabdodontomorpha. Однако, размещение Burianosaurus в этой кладе следует рассматривать с осторожность, поскольку оригинальное исследование Макдональда не было сосредоточено на изучении взаимоотношений таксонов за пределами Ankylopollexia. Авторы считают, что такой результат в большей степени опровергает первоначальное отнесение Burianosaurus к игуанодонтидам.
Второй набор данных основывался на исследовании Клинта Бойда 2015 года[5]. Целью включения Burianosaurus в этот анализ было выяснение филогенетического положения динозавра среди орнитопод вне клады Ankylopollexia, а конкретнее, выяснение отношений с европейскими дриозавридами и Rhabdodontomorpha. В этом наборе данных закодировано уже 14 признаков Burianosaurus. На дереве строго консенсуса, полученном из 18133 максимально экономных деревьев, Burianosaurus находится за пределами клады Iguanodontia (в определении Бойда, 2015) в политомии с гондванской кладой Elasmaria. Политомия объясняется фрагментарным материалом члена этой клады — Morrosaurus. Во всех экономных деревьях Burianosaurus находится чуть ближе к гадрозавридам, чем Elasmaria, но базальнее последнего общего предка Iguanodontia, а положение Morrosaurus меняется от сестринского к Burianosaurus и Iguanodontia, до сестринского к Iguanodontia внутри Elasmaria. После удаления Morrosaurus был проведён второй анализ. На дереве строго консенсуса, полученном из 13547 максимально экономных деревьев, Burianosaurus занимает сестринское положение к Iguanodontia. Эта клада характеризуется одним признаком: четвёртый вертел находится на середине тела кости или ниже (присутствует также у Thescelosaurus neglectus, Thescelosaurus assiniboiensis и Parksosaurus; отсутствует у нескольких представителей Rhabdodontomorpha и у всех дриозаврид).
Кладограмма строго консенсуса на основе модифицированной матрицы данных Boyd, 2015:
| Clypeodonta |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Примечания
- Madzia, Daniel; Boyd, Clint A.; Mazuch, Martin (2017). «A basal ornithopod dinosaur from the Cenomanian of the Czech Republic». Journal of Systematic Palaeontology: 1-13.
- Fejfar, O.; Kostak, M.; Kvacek, J.; Mazuch, M.; Moucka, M. (2005). «First Cenomanian dinosaur from Central Europe (Czech Republic)». Acta Palaeontologica Polonica. 50 (2): 295—300.
- Andrew T. McDonald (2012). «Phylogeny of Basal Iguanodonts (Dinosauria: Ornithischia): An Update». PLoS ONE. 7 (5): e36745
- Boyd, C. A. & Pagnac, D. C. 2015. «Insight on the anatomy, systematic relationships, and age of the Early Cretaceous ankylopollexian dinosaur Dakotadon lakotaensis». PeerJ, 3, e1263.
- Boyd, C. A. 2015. «The systematic relationships and biogeographic history of ornithischian dinosaurs». PeerJ, 3, e1523.