Эксперимент Мезельсона и Сталя
Экспериме́нт Мезельсо́на и Ста́ля (англ. Meselson–Stahl experiment — эксперимент, проведённый двумя молекулярными биологами — Мэтью Мезельсоном и Франклином Сталем в 1958 году. Он показал, что репликация ДНК имеет полуконсервативный характер[1]. Это означает, что каждая дочерняя двойная спираль ДНК состоит из одной старой (матричной) цепи и из одной вновь синтезированной цепи.
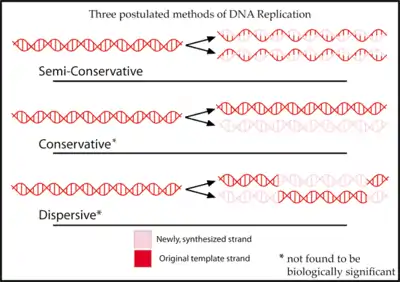
Предварительные гипотезы
После открытия Уотсоном и Криком двойной спирали ДНК было предложено несколько возможных механизмов её репликации. Первую гипотезу полуконсервативной репликации ДНК предложили сами Уотсон и Крик[2].
Гипотеза консервативной репликации ДНК предполагает, что материнская двойная спираль как целое выступает в качестве матрицы для синтеза дочерней спирали, состоящей из двух новых цепочек[3]. Эта гипотеза подразумевает большую роль гистонов в процессе репликации.
Гипотеза дисперсной репликации возникла как попытка объяснить, каким образом клетка может решить проблему раскручивания длинных дуплексов при копировании ДНК. Согласно этой гипотезе, для предотвращения суперскручивания ДНК при репликации в неё через каждые 5 нуклеотидных остатков вносятся разрывы, которые «зашиваются» после того, как излишнее напряжение снимется с молекулы. В результате дочерняя (вновь синтезируемая цепь) состоит из чередующихся старых и новых участков длиной по 5 нуклеотидных остатков. То же верно и для материнской цепи. Эта гипотеза была предложена Максом Дельбрюком[4].
Каждая из этих гипотез предполагает определённое распределение старой ДНК в молекулах, образующихся после завершения репликации. По гипотезе консервативной репликации одна из молекул будет полностью старой, а вторая — полностью новой. Полуконсервативный синтез должен приводить к формированию молекул, которые содержат по одной старой и одной новой цепи. Модель дисперсной репликации же предсказывает, что каждая цепь каждой молекулы ДНК будет состоять из чередующихся старых и новых участков[5]. Таким образом, если установить, какой из этих случаев наблюдается в природе, можно определить верную модель.
Схема эксперимента и результаты
В 1957 году Мезельсон, Сталь и Джером Виноград опубликовали статью о новом методе изучения молекулярного веса и парциального удельного объёма макромолекул (например, ДНК) — равновесном ультрацентрифугировании в градиенте плотности[6]. Этот метод позволяет разделять молекулы ДНК по их плотности: каждая молекула остановится в том месте градиента, где плотность раствора совпадает с её плавучей плотностью. Авторы применили этот метод для разделения молекул ДНК, содержащих изотопы азота 14N и 15N[1]. 15N не радиоактивен, а лишь тяжелее 14N. Содержащие тяжелый изотоп молекулы ДНК функциональны и могут удваиваться.
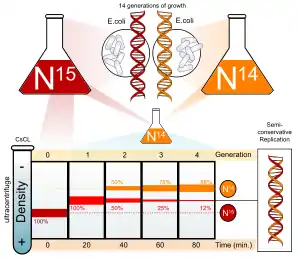
Мезельсон и Сталь показали, что, если вырастить несколько поколений бактерий Escherichia coli в среде, богатой 15N или 14N, затем центрифугировать их ДНК в градиенте плотности хлористого цезия, то окажется, что более тяжёлая 15N-ДНК останавливается ближе ко дну центрифужной пробирки, чем 14N-ДНК[1].
Для того чтобы установить механизм репликации, E. coli, которые в течение нескольких поколений росли в 15N-содержащей среде (а значит, их ДНК содержала только 15N) были перенесены в 14N-содержащую среду, где им было позволено поделиться только один раз. Плотность выделенной из этих клеток ДНК оказалась больше плотности ДНК бактерий, выращенных в среде, богатой 14N, но меньше плотности ДНК бактерий, выращенных в 15N среде. Это противоречило гипотезе о консервативном характере репликации ДНК, при котором ДНК разделились бы на две фракции с высокой и низкой плотностью, но не с промежуточной. Таким образом, первая гипотеза была отброшена[1].
Однако полученный результат не исключал дисперсный механизм репликации, при котором участки материнской ДНК чередуются с участками дочерней ДНК. По гипотезе дисперсной репликации плотность ДНК бактерий должна быть одинаковой для всех молекул и занимать промежуточное положение между плотностью ДНК клеток первого поколения и плотностью самой лёгкой ДНК. Оказалось, однако, что клетки содержали примерно равные количества тяжелых ДНК (первое поколение) и гибридных ДНК (второе поколение). Этот факт позволил исключить гипотезу дисперсного механизма репликации[1].
Примечания
- Meselson M., Stahl F. W. THE REPLICATION OF DNA IN ESCHERICHIA COLI. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1958. — 15 July (vol. 44, no. 7). — P. 671—682. — PMID 16590258.
- WATSON JD, CRICK FH. Genetical implications of the structure of deoxyribonucleic acid. (англ.) // Nature. — 1953. — 30 May (vol. 171, no. 4361). — P. 964—967. — PMID 13063483.
- Bloch D. P. A POSSIBLE MECHANISM FOR THE REPLICATION OF THE HELICAL STRUCTURE OF DESOXYRIBONUCLEIC ACID. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1955. — 15 December (vol. 41, no. 12). — P. 1058—1064. — PMID 16589796.
- Delbrück M. ON THE REPLICATION OF DESOXYRIBONUCLEIC ACID (DNA). (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1954. — September (vol. 40, no. 9). — P. 783—788. — PMID 16589559.
- M. Delbrück, G. S.Stent. On the mechanism of DNA replication // A Symposium on the Chemical Basis of Heredity (англ.) / McElroy, William D.; Glass, Bentley. — Johns Hopkins Pr., 1957. — P. 699—736.
- Meselson M., Stahl F. W., Vinograd J. EQUILIBRIUM SEDIMENTATION OF MACROMOLECULES IN DENSITY GRADIENTS. (англ.) // Proceedings Of The National Academy Of Sciences Of The United States Of America. — 1957. — 15 July (vol. 43, no. 7). — P. 581—588. — PMID 16590059.