Флоэма
Флоэ́ма (от греч. φλοῦς — кора) — проводящая ткань сосудистых растений, по которой происходит транспорт продуктов фотосинтеза к частям растения, в которых он не происходит: подземные части, конусы нарастания, цветки, плоды и др. Вместе с ксилемой (древесиной), обеспечивающей транспорт воды и минеральных солей, образует проводящие пучки.
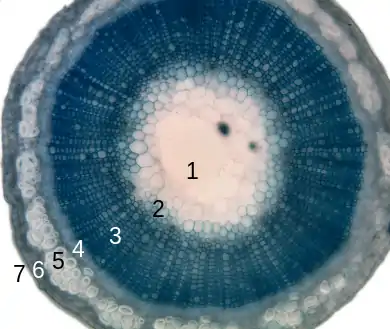
1. рыхлая сердцевина,
2. протоксилема,
3. ксилема,
4. флоэма,
5. склеренхима,
6. перидерма,
7. эпидерма
Морфология
В стеблях большинства растений флоэма располагается снаружи по отношению к ксилеме, a в листьях — обращена к нижней стороне жилок листовой пластинки, в проводящих пучках корней тяжи флоэмы и ксилемы чередуются.
По своему происхождению флоэма делится на первичную, дифференциирующуюся из прокамбия и вторичную, дифференциирующуюся из камбия. Первичная флоэма, в свою очередь, подразделяется на протофлоэму и метафлоэму, в отличие от вторичной флоэмы, у первичной отсутствуют сердцевинные лучи.
Клеточный состав и первичной, и вторичной флоэмы одинаков; они состоят из нескольких типов клеток различной морфологии, выполняющих различные функции:
- Ситовидные элементы (ситовидные клетки, ситовидные трубки и клетки-спутницы), обеспечивающие основной транспорт
- Склеренхимные элементы (склереиды и волокна), выполняющие опорную функцию
- Паренхимные элементы (паренхимные клетки), обеспечивающие ближний радиальный транспорт
Ситовидные элементы являются высокоспециализированными клетками, обеспечивающими флоэмный транспорт ассимилятов. Их особенностью, благодаря которой они получили своё название, являются ситовидные поля (или ситовидные пластинки у покрытосеменных растений), являющихся скоплением специализированных пор в клеточной стенке. Поры ситовидных полей являются видоизменёнными первичными поровыми полями — их поры представляют собой расширенные плазмодесмы — цитоплазматические мостики, соединяющие соседние протопласты, однако если первичное поровое поле обычно пронизано несколькими плазмодесмами, через пору ситовидного поля проходит один цитоплазматический тяж с диаметром до нескольких микрометров, что в десятки раз превышает диаметр плазмодесм. Происхождение такого гигантского по сравнению с плазмодесмами цитоплазматического тяжа неясно, считается, что его образование возможно как путём слияния группы плазмодесм, так расширением единственной плазмодесмы.
Канал поры ситовидного элемента выстлан полисахаридом каллозой, которая у многих растений образует валик у отверстия канала, каллоза накапливается в течение жизни ситовидного элемента.
Ситовидные элементы споровых (хвощей, плаунов, папоротников) и голосеменных растений представлены ситовидными клетками, эти клетки вытянуты в длину, ситовидные поля разбросаны по боковым стенкам.
Флоэмный транспорт
Флоэмный сок представляет собой раствор углеводов (у древесных растений — преимущественно сахарозы), являющихся продуктами фотосинтеза, в достаточно высокой концентрации — 0,2—0,7 моль/литр (7—25 %), а также других ассимилятов и метаболитов (аминокислот и фитогормонов) в значительно более низких концентрациях. Скорость транспорта составляет десятки сантиметров в час, что превышает скорость диффузии.
В отличие от ксилемы, где перемещение содержимого происходит в одном направлении — вверх от корней к листьям, флоэмный транспорт происходит от органов-доноров, в которых происходит фотосинтез к акцепторам — органам или областям, в которых продукты фотосинтеза потребляются или запасаются. Интенсивное потребление ассимилятов происходит в корнях, верхушках побегов, формирующихся листьях, репродуктивных органах; у многих видов растений есть специальные органы запасания — луковицы, клубни и корневища, выступающие в качестве акцепторов.
Эксперименты с радиоактивными метками (14C) показали, что транспорт от донора — например, подкормленного меткой листа — происходит к ближайшим акцепторам, то есть нижние листья обеспечивают углеводами корни, листья побега, расположенные рядом с плодами — плоды и т. п. Следует отметить, что флоэмный транспорт является двусторонним: так, органы запасания могут в зависимости от фазы вегетации выступать как в качестве акцептора — при накоплении крахмала, синтезирующегося из углеводов, поставляемых листьями в конце вегетационного периода — так и в качестве донора при расщеплении запасённого крахмала на углеводы, идущие на построение молодых листьев в начале периода вегетации.
Литература
- Гэлстон А., Девис П., Сэттер Р. Жизнь зелёного растения. — М., Мир, 1983.
Ссылки
- Флоэма — статья из Большой советской энциклопедии.