Униониды
Униони́ды[1] или перло́вицы[2][3] (лат. Unionidae), — семейство пресноводных моллюсков из отряда унионид (Unionida)[4]. Самое большое семейство в составе класса двустворчатых[5]: включает около 40 родов[4] и, по разным оценкам, около 1000 видов[6]. К наиболее известным представителям семейства можно отнести беззубок (Anodonta) и перловиц (Unio)[7].
| Униониды | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Раковина Anodonta anatina | ||||||||||||
| Научная классификация | ||||||||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Без ранга: Тип: Класс: Подкласс: Отряд: Семейство: Униониды |
||||||||||||
| Международное научное название | ||||||||||||
| Unionidae Rafinesque, 1820 | ||||||||||||
| Роды | ||||||||||||
| ||||||||||||
Характерной особенностью семейства является развитие с особой личиночной стадией, паразитирующей на рыбах — глохидием, отсутствие ноги у взрослых форм и слабое развитие сифонов. Для унионид характерен уникальный механизм наследования митохондриальной ДНК.
Многие представители семейства употребляются в пищу животными и человеком; их раковины также находят применение благодаря красивому перламутру[8]. Многие виды унионид находятся под угрозой исчезновения и включены в Красные книги МСОП и России.
Строение
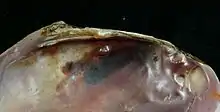
Раковина длиной от 3 до 25 см[6], с равными створками, изнутри покрытыми хорошо развитым перламутровым слоем. Лигамент наружный. Замковый аппарат гетеродонтный, с небольшим количеством зубов или отсутствует.
Иногда замковый аппарат унионид описывают как «псевдогетеродонтный», или унионидный: зубы могут быть более или менее расщеплёнными, расходящимися и несут функции центральных или боковых зубьев.
Нога клиновидная. У взрослых форм биссусная железа отсутствует, однако она имеется у личинок. Жабры пластинчатые[8]. Сифоны обычно развиты слабо[9], края листков мантии несросшиеся[10]. У многих представителей подсемейства лампсилин (Lampsilinae) выражен половой диморфизм, проявляющийся даже в форме их раковины[11].
Размножение и развитие
Глохидии
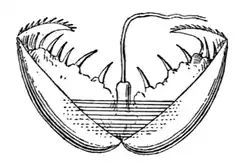

Униониды известны своим характерным сложным жизненным циклом. Практически все виды, за исключением отдельных гермафродитных, например, Elliptio complanata[12] раздельнополые.
Сперма выводится из мантийной полости мужской особи через выводящий проток и попадает в женскую мантийную полость через соответствующее вводное отверстие. Оплодотворённые яйца переносятся из гонад в жабры, где молодые особи вылупляются и достигают личиночной стадии — глохидия. Развитие молоди до стадии глохидия происходит внутри жаберных полостей, играющих роль выводковых сумок. Сформировавшиеся глохидии имеют двустворчатую раковину с округлыми створками, подостланную личиночной мантией, которая несёт по краю чувствительные щетинки. У них имеется зачаточные нога, жабры и кишка. Глохидий может плавать, хлопая створками, приводимыми в движение единственным мускулом-замыкателем[13]. Рта, ануса и развитого пищеварительного тракта у глохидиев нет, но у многих видов они снабжены биссусовой нитью. Размер глохидиев варьирует от 50 до 500 мкм в зависимости от видовой принадлежности[14].
Глохидии выводятся из тела самки через сифон в окружающую среду, причём способ выбрасывания зависит от образа жизни рыбы-хозяина. Некоторые моллюски формируют на дне слизистые тяжи, в которые заключены личинки; ими заражаются рыбы, нерестящиеся на дне или питающиеся бентосными организмами. У некоторых унионид глохидии образуют плотные окрашенные скопления, имитирующие пищу рыб-хозяев: червей, насекомых или мальков, и таким образом привлекают рыб. Некоторые униониды развили весьма необычную стратегию размножения. Мантия самки выдаётся из раковины и имитирует маленькую рыбу: на ней появляются отметины, напоминающие чешую рыб, и ложные глаза. Эта приманка привлекает внимание настоящих рыб. Некоторые рыбы рассматривают её как добычу, другие видят в ней особь своего вида. Рыбы подплывают ближе, чтобы рассмотреть приманку, и тогда моллюск выбрасывает прямо на рыбу струю воды, содержащую целое облако глохидиев, которые внедряются в её жабры или покровы[14].
Прикрепление глохидия осуществляется при помощи биссуса. Если у глохидиев имеются, кроме биссуса, крючковидные выросты на створках, то они прикрепляются к чешуе рыбы, причём при контакте с рыбой створки глохидия немедленно захлопываются, а если этих выростов нет, то глохидий прикрепляется к жабрам, при этом доставка глохидиев к жабрам осуществляется током воды. Сигналом для смыкания створок служат особые вещества, содержащиеся в слизи на покровах рыбы. После прикрепления к рыбе личинку начинает затягивать её эпителий, и вокруг неё формируется циста, состоящая из эпителиальной и соединительной тканей рыбы. В мантии личинки присутствуют фагоцитарные клетки, которые разрушают ткани рыбы и поглощают образовавшийся органический материал, обеспечивая тем самым питание глохидия. В этой цисте глохидий проводит от 10 до 30 дней, а иногда и несколько месяцев, при этом личиночные органы глохидия дегенерируют и заменяются органами, присущими взрослой особи. В конце концов молодой моллюск разрушает стенку цисты, падает на дно и закапывается в грунт, и в дальнейшем он постепенно переходит к образу жизни, свойственному взрослым особям[15].
Для некоторых унионид характерна узкая специфичность в выборе хозяина, однако большая часть унионид использует не один, а несколько видов рыб-хозяев, и на одном и том же виде рыб могут развиваться глохидии нескольких видов моллюсков[14]. При прикреплении глохидия к резистентной рыбе или рыбе не вида-хозяина происходят сбои в перестройке эпителия в месте прикрепления личинки, и такие нарушения в формировании цисты могут снизить эффективность метаморфоза или вовсе привести к гибели личинки[16].
Таким образом, глохидий является эктопаразитом, а рыбы обеспечивают его распространение[13]. Крупные униониды могут ежегодно продуцировать до 17 миллионов глохидиев, при этом на одной особи рыбы могут одновременно развиваться до 3000 личинок. Впрочем, серьёзного ущерба взрослой рыбе глохидии обычно не приносят, и тем не менее мальки часто погибают от вторичной инфекции[17]. В ходе недавних исследований было показано, что глохидии могут изменять поведение рыб-хозяев, делая их более пассивными и вызывая небольшие изменения среды обитания, однако эти изменения поведения не направлены на увеличение расселения моллюсков[18]. Униониды и рыбы-хозяева их глохидиев тесно связаны друг с другом. В частности, была показана линейная связь между количеством видов рыб и видов унионид в одном водоёме, и изменения в видовом составе рыб водоёма приводят к изменению видового состава унионид в этом водоёме[19].
Генетика
У унионид наблюдается уникальный механизм наследования митохондриальной ДНК, причём мтДНК также вовлечена в определение пола. Пол унионид определяется нуклеотидной последовательностью в составе митохондриальной ДНК — мужской (M-ORF) или женской (F-ORF) открытой рамкой считывания; таким образом, у них происходит наследование мтДНК как по мужской, так и по женской линии. Представители гермафродитных видов лишены обеих последовательностей, однако несут напоминающую F-ORF гермафродитную открытую рамку считывания (H-ORF). Предполагается, что для предковых форм большинства моллюсков был характерен гермафродитизм с дальнейшим появлением отдельно мужских особей. Кроме того, участок м-ДНК, определяющий пол, мог сыграть ключевую роль в эволюции двойного унипарентального наследования, характерного для некоторых пресноводных моллюсков[20]. Наконец, анализ женских и мужских митохондриальных последовательностей используется в филогенетических исследованиях[21].
Распространение
Распространены практически повсеместно; как правило, встречаются в лотических экосистемах (пресноводные водоемы с проточной водой). Наибольшее известное число таксонов (297) насчитывается в Северной Америке[22][23][24], однако территории Китая и Юго-Восточной Азии также отличаются разнообразием унионид[25][26]. В реках европейской части России наиболее часто встречаются беззубки, в реках Дальнего Востока нередки гребенчатки (Cristaria)[8], а наибольшее разнообразие перловиц наблюдается в Восточной Азии[26].
Экология
Униониды закапываются в субстрат створками наружу. Они являются фильтраторами, отфильтровывают из воды фито- и зоопланктон, бактерии, грибные споры и мертвую органику[27][28][29][30][31][32][33][34][35][36]. Несмотря на многочисленные исследования, конечный продукт обмена моллюсков не ясен. При высокой плотности взвешенной органики моллюски регулируют уровень фильтрации и, соответственно, прозрачность воды[37][38], однако объём и скорость фильтрации зависят от внешних факторов (температура воды, скорость течения, размер и концентрация фильтрата). Кроме того, объём фильтрации и размер захватываемых частиц зависит от морфологии жабр[31]. Таким образом, они выступают в роли природных биофильтров и регулируют самоочищение воды; кроме того, многие из них являются индикаторами состояния водной среды[39].
Палеонтология
В больших количествах раковины двустворчатых моллюсков могут влиять на состояние других ископаемых останков. Например, два единственных известных микрофрагмента скорлупы яиц гадрозавров были обнаружены в свите Dinosaur Park Formation, где сохранились исключительно благодаря наличию вокруг раковин древних беспозвоночных, в том числе и унионид. Как правило, скорлупа разрушается под действием таннинов окружающих хвойных, уменьшающих pH грунтовых вод. Однако медленное разложение известковых раковин моллюсков сопровождается выделением карбоната кальция, что восстанавливает нейтральность среды и создает условия для фоссилизации скорлупы[40].
Охрана

По состоянию на август 2014 года в Международной Красной книге МСОП имеется информация о 438 видах унионид, при этом 131 вид находится в разной степени риска (категории CR, EN, VU), а 28 видов признаны вымершими[42]. В Красную книгу России занесены следующие 13 видов унионид: 7 видов рода миддендорффиная (Middendorffinaia), 1 вид рода нодулярия (Nodularia), 4 вида рода ланцеолярия (Lanceolaria), 1 вид рода гребенчатки (Cristaria)[43].
Для двустворчатых, находящихся под угрозой исчезновения, характерен ряд общих черт: позднее половое созревание, относительно высокая продолжительность жизни, низкая плодовитость, ограниченный ареал, специфическая среда обитания, а для унионид дополнительным фактором является наличие специфических хозяев для глохидиев[44].
Хозяйственное значение и применение
Многие представители семейства унионид нашли разнообразное практическое применение. В частности, в некоторых странах унионид используют в пищу, особенно для откорма животных. Раньше из раковин унионид делали бельевые пуговицы, имеются промысловые виды перловиц и беззубок с красивым перламутром. К их числу относятся гребенчатки, раковина которых может достигать 34 см. В Китае, Японии и Индокитае из гребенчаток изготавливают перламутровые изделия[8]. В США вылавливают и разводят представителей подсемейства лампсилин (Lampsilinae), особенно некоторые быстрорастущие виды из рода Lampsilis и других. При искусственном разведении моллюсков рыб специально заражают их глохидиями [11].
Классификация
Некоторые молекулярно-филогенетические исследования указывали на парафилетичность унионид, что, однако, расходилось с морфологическими данными. В молекулярном исследовании 2011 года униониды были признаны группой с надёжно установленной монофилией[45].
Систематика унионид является одной из самых запутанных и неопределённых среди двустворчатых моллюсков. Это обусловлено значительной изменчивостью раковин унионид: раковины взрослых моллюсков, обычно используемые в классификации, не имеют выраженных видоспецифичных признаков (особенно это касается беззубок, чьи раковины не имеют замка). По этой причине имеются разногласия в выделении видов, родов и даже подсемейств[39]. Как упоминалось выше, в настоящее время в классификации, помимо морфологических признаков, также используется анализ митохондриальных геномов унионид. Согласно наиболее общепринятым современным представлениям, в семейства Униониды насчитывается 43 рода[4]:
|
|
 Actinonaias pectorosa
Actinonaias pectorosa Amblema plicata
Amblema plicata Elliptio steinstansana
Elliptio steinstansana Fusconaia subrotunda
Fusconaia subrotunda Lampsilis abrupta
Lampsilis abrupta
Примечания
- Аннотированный перечень редких и находящихся под угрозой исчезновения видов беспозвоночных животных, особо охраняемых в пределах России // 2003* Россия* Красный список особо охраняемых редких и находящихся под угрозой исчезновения животных и растений. (2-й выпуск). — Часть 2: Беспозвоночные животные (Бюллетень Красной книги, 2/2004 (2008)) / отв. ред. В. Е. Присяжнюк. — М.: Лаборатория Красной книги Всероссийского научно-исследовательского института охраны природы, 2004 (2008). — 512 с. — ISBN 978-5-9243-0158-7. — С. 94. Полный текст Архивная копия от 24 октября 2018 на Wayback Machine
- Перловицы / Чесунов А. В. // П — Пертурбационная функция. — М. : Большая российская энциклопедия, 2014. — С. 701. — (Большая российская энциклопедия : [в 35 т.] / гл. ред. Ю. С. Осипов ; 2004—2017, т. 25). — ISBN 978-5-85270-362-0.
- Жизнь животных, 1988, с. 83.
- Семейство Unionidae (англ.) в Мировом реестре морских видов (World Register of Marine Species) (Дата обращения: 9 августа 2014).
- Huber, Markus. Compendium of Bivalves. A Full-color Guide to 3'300 of the World's Marine Bivalves. A Status on Bivalvia after 250 Years of Research (англ.). — Hackenheim: ConchBooks, 2010. — P. 901 pp. + CD. — ISBN 978-3-939767-28-2.
- Animal Diversity Web: Unionidae.
- Жизнь животных, 1968, с. 134—135.
- Шарова, 2002, с. 314.
- Рупперт, Фокс, Барнс, 2008, с. 260.
- Жизнь животных, 1968, с. 132.
- Жизнь животных, 1968, с. 136.
- Downing J. A., Amyot J. P., Pérusse M., Rochon Y. Visceral sex, hermaphroditism, and protandry in a population of the freshwater bivalve Elliptio complanata // Journal of the North American Benthological Society. — Vol. 8, № 1. — P. 92—99.
- Шарова, 2002, с. 312.
- Рупперт, Фокс, Барнс, 2008, с. 258.
- Рупперт, Фокс, Барнс, 2008, с. 258—259.
- Rogers-Lowery C. L., Dimock R. V. Jr. Encapsulation of attached ectoparasitic glochidia larvae of freshwater mussels by epithelial tissue on fins of naive and resistant host fish. (англ.) // The Biological bulletin. — 2006. — Vol. 210, no. 1. — P. 51—63. — PMID 16501064.
- Рупперт, Фокс, Барнс, 2008, с. 259.
- Pavel Horký, Karel Douda, Matúš Maciak, Libor Závorka, Ondřej Slavík. Parasite-induced alterations of host behaviour in a riverine fish: the effects of glochidia on host dispersal // Freshwater Biology. — 2014. — Vol. 59, № 7. — P. 1452—1461. — doi:10.1111/fwb.12357.
- J. A. Daraio, L. J. Weber, Steven J. Zigler, Teresa J. Newton, John M. Nestler. Simulated effects of host fish distirbution on juvenile unionid mussels dispersal in a large river // RIVER RESEARCH AND APPLICATIONS. — 2012. — Vol. 28. — P. 594—608. — doi:10.1002/rra.1469.
- Breton S., Stewart D. T., Shepardson S., Trdan R. J., Bogan A. E., Chapman E. G., Ruminas A. J., Piontkivska H., Hoeh W. R. Novel protein genes in animal mtDNA: a new sex determination system in freshwater mussels (Bivalvia: Unionoida)? (англ.) // Molecular biology and evolution. — 2011. — Vol. 28, no. 5. — P. 1645—1659. — doi:10.1093/molbev/msq345. — PMID 21172831.
- Doucet-Beaupré H., Blier P. U., Chapman E. G., Piontkivska H., Dufresne F., Sietman B. E., Mulcrone R. S., Hoeh W. R. Pyganodon (Bivalvia: Unionoida: Unionidae) phylogenetics: a male- and female-transmitted mitochondrial DNA perspective. (англ.) // Molecular phylogenetics and evolution. — 2012. — Vol. 63, no. 2. — P. 430—444. — doi:10.1016/j.ympev.2012.01.017. — PMID 22326838.
- Williams, J. D., M. L. Warren, K. S. Cummings, J. L. Harris, and R. J. Neves. Conservation Status of Freshwater Mussels of the United States and Canada (англ.) // Fisheries : journal. — 1993. — Vol. 18, no. 9. — P. 6—22. — ISSN 1548-8446. — doi:10.1577/1548-8446(1993)018<0006:CSOFMO>2.0.CO;2.
- Burch, John B. Freshwater unionacean clams (Mollusca: Pelecypoda) of North America. — 1973. — P. 114. — 176 p.
- Heard, William H. Identification Manual of the Freshwater Clams of Florida. — 1979. — Vol. 4. — P. 1—83.
- Freshwater Molluscan Shells: Unionidae.
- Жизнь животных, 1968, с. 134.
- Allan W. R. The food and feeding habits of freshwater mussels // Biological Bulletin. — 1914. — Vol. 27. — P. 127—147.
- Coker R. E., Shira A.F., Clark H. W., Howard, A. D. Natural history and propagation of fresh-water mussels // Bulletin of the Bureau of Fisheries. — 1921. — Vol. 37. — P. 77—181.
- Churchill E. P., Lewis S. I. Food and feeding in fresh-water mussels // Bulletin of the Bureau of Fisheries. — 1924. — Vol. 39. — P. 439—471.
- James H. Thorp, Alan P. Covich. Ecology and classification of North American freshwater invertebrates. — San Diego: Academic Press, 2001. — P. 331—429. — 1056 p.
- Silverman H., Nichols S. J., Cherry J. S., Archberger E., Lynn J. S., Dietz T. H. Clearance of laboratory-cultured bacteria by freshwater bivalves: differences between lentic and lotic unionids // Canadian Journal of Zoology. — 1997. — Vol. 75. — P. 1857—1866.
- Bärlocher F., Brendelberger H. Clearance of aquatic hyphomycete spores by a benthic suspension feeder // Limnology and Oceanography. — 2004. — Vol. 49, № 6. — P. 2292—2296. — doi:10.4319/lo.2004.49.6.2292. Архивировано 23 сентября 2015 года.
- Roditi H. A., Fisher N. S., Sañudo-Wilhelmy S. A. Uptake of dissolved organic carbon and trace elements by zebra mussels. (англ.) // Nature. — 2000. — Vol. 407, no. 6800. — P. 78—80. — doi:10.1038/35024069. — PMID 10993076.
- Baines S. B., Fisher N. S., Cole J. J. Uptake of dissolved organic matter (DOM) and its importance to metabolic requirements of the zebra mussel, Dreissena polymorpha // Limnology and Oceanography. — 2005. — Vol. 50, № 1. — P. 36—47.
- Yeager M. M., Cherry D. S., Neves R. J. Feeding and burrowing behaviors of juvenile rainbow mussels, Villosa iris (Bivalvia, Unionidae) // Journal of the North American Benthological Society. — 1994. — Vol. 13, № 2. — P. 217—222.
- Nichols S. J., Silverman H., Dietx T. H., Lynn J. W., Garling D. L. Pathways of food uptake in native (Unionidae) and introduced (Corbiculidae and Dreissenidae) freshwater bivalves // Journal of Great Lakes Research. — 2005. — Vol. 31, № 1. — P. 87—96. — doi:10.1016/S0380-1330(05)70240-9.
- Cohen R. R. H., Dresler P. V., Phillips E. P. J., Cory R. L. The effects of the Asiatic clam, Corbicula fluminea, on phytoplankton of the Potomac River, Maryland // Limnology and Oceanography. — 1984. — Vol. 29, № 1. — P. 170—180. Архивировано 23 сентября 2015 года.
- Phelps H. L. The Asiatic clam (Corbicula fluminea): invasion and system-level ecological change in the Potomac River estuary near Washington, D. C // Estuaries. — 1994. — Vol. 17, № 3. — P. 614—621.
- Саенко Е. М. Беззубки (Bivalvia: Unionidae: Anodontinae) российского Дальнего Востока. — 2003. (недоступная ссылка)
- Tanke D. H., Brett-Surman M. K. Evidence of Hatchling and Nestling-Size Hadrosaurs (Reptilia:Ornithischia) from Dinosaur Provincial Park (Dinosaur Park Formation: Campanian), Alberta, Canada / D. H. Tanke, K. Carpenter. — Mesozoic Vertebrate Life—New Research Inspired by the Paleontology of Philip J. Currie.. — Bloomington: Indiana University Press, 2011. — P. 206—218. — xviii + 577 p.
- Alasmidonta raveneliana (англ.). The IUCN Red List of Threatened Species.
- IUCN Red List of Threatened Species. Version 2014.2.
- Красная книга России: моллюски. Дата обращения: 19 августа 2013.
- The Conservation Biology of Molluscs // E. Alison Kay. — International Union for Conservation of Nature and Natural Resources, 1995. — ISBN 2-8317-0053-1.
- Whelan N. V., Geneva A. J., Graf D. L. Molecular phylogenetic analysis of tropical freshwater mussels (Mollusca: Bivalvia: Unionoida) resolves the position of Coelatura and supports a monophyletic Unionidae. (англ.) // Molecular phylogenetics and evolution. — 2011. — Vol. 61, no. 2. — P. 504—514. — doi:10.1016/j.ympev.2011.07.016. — PMID 21827862.
- Жизнь животных, 1988, с. 84.
Литература
- Жадин В. И. Сем. Unionidae. — Фауна СССР. 18. — М.—Л., 1938.
- Жизнь животных. В 6 т. / гл. ред. Л. А. Зенкевич. — 1‑е изд. — М. : Просвещение, 1968. — Т. 2 : Беспозвоночные / под ред. Л. А. Зенкевича. — 563 с. : ил. — 300 000 экз.
- Жизнь животных. В 7 т. / гл. ред. В. Е. Соколов. — 2‑е изд., перераб. — М. : Просвещение, 1988. — Т. 2 : Моллюски. Иглокожие. Погонофоры. Щетинкочелюстные. Полухордовые. Хордовые. Членистоногие. Ракообразные / под ред. Р. К. Пастернак. — 447 с. : ил. — ISBN 5-09-000445-5.
- Рупперт Э. Э., Фокс Р. С., Барнс Р. Д. Зоология беспозвоночных: функциональные и эволюционные аспекты. — М.: Издательский центр «Академия», 2008. — Т. 2. — 448 с. — ISBN 978-5-7695-3495-9.
- Шарова И. Х. Зоология беспозвоночных. — М.: Владос, 2002. — 592 с. — ISBN 5-691-00332-1.
- Догель В. А. Зоология беспозвоночных. — 7-е изд. — М.: Высшая школа, 1981. — 614 с.
Ссылки
- Missouri State Unio Gallery.
- Freshwater Mollusk Conservation Society.
- Ohio State University: Division of Molluscs — Freshwater Mussel Collection — Unionidae.
- Unionidae at The MUSSEL Project Web Site.
| Таксономия |
|---|