Глубоководные рыбы
Глубоководные рыбы — рыбы, которые населяют склон и ложе океана, а также толщу воды от нижней границы эпипелагиали до абиссали. Количество видов глубоководных рыб достигает 2000, на глубине свыше 6000 м обитает не более 10—15 видов. По типу питания глубоководные рыбы делятся на планктонофагов, бентофагов и хищников. Кроме того, их можно в целом разделить на 2 группы: истинно глубоководных, имеющих специфические органы (телескопические глаза, органы свечения и т. д.) и шельфоглубоководных, не имеющих подобных адаптаций и населяющих материковый склон. Наибольшее биоразнообразие наблюдается у рыб, обитающих в верхних слоях батиали (200—3000 м). В крупных озёрах, за исключением Байкала, глубоководные рыбы отсутствуют[1].

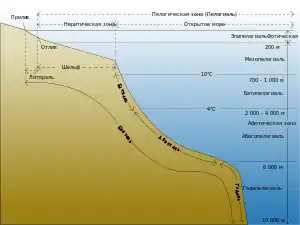
Глубоководные рыбы обитают в эвфотической зоне, лежащей под залитой солнцем пелагиалью. Самой распространённой глубоководной рыбой является светящийся анчоус. Также на глубине встречаются фонареглазовые, бразильские светящиеся акулы, гоностомовые, удильщики, хаулиоды и некоторые виды бельдюг. В глубоководной зоне довольно многочисленны и активны донные рыбы[2]. Здесь распространены долгохвостовые, бититовые, бельдюги, лопаточные нетопыри и пинагоры[3].
Лишь 2 % известных морских рыб обитают в пелагиали[4]. Глубоководные организмы в целом населяют батипелагическую (1000—4000 м) и абиссопелагическую (4000—6000 м) афотические зоны. Именно эти зоны составляют около 75 % от обитаемого пространства Мирового океана. В зависимости от района в океане на глубине 700—1000 м пролегает слой с минимальным содержанием кислорода. Эта зона наиболее богата питательными веществами. Свойства, характерные для глубоководных организмов, например, биолюминесценция, встречаются и у обитателей мезопелагиали (200—1000 м). Мезопелагиаль — это сумеречная зона, в которую проникает очень мало солнечного света, но он всё же есть[5].
Эпипелагиаль (0—200 м) — фотическая зона, в которую проникает солнечный свет, здесь протекает фотосинтез. Однако 90 % объёма Мирового океана погружено во тьму, температура воды здесь не превышает 3 °C и понижается до −1,8 °C (за исключением гидротермальных экосистем, где температура превосходит 350 °C), мало кислорода, а давление колеблется в пределах 20—1000 атмосфер[6].
Окружающая среда

За краем континентального шельфа постепенно начинаются абиссальные глубины. Здесь проходит граница между прибрежными, довольно мелкими бентическими зонами обитания и глубоководной бенталью. Площадь этой пограничной территории составляет около 28 % площади Мирового океана[7].
Под эпипелагиалью находится обширная водная толща, в которой обитают разнообразные организмы, приспособленные к условиям жизни на глубине[8]. На глубине между 200 и 1000 м освещённость ослабевает до тех пор, пока не наступает полная темнота. Через термоклин температура падает до 4—8 °C. Это сумеречная или мезопелагическая зона.
Около 40 % дна океана состоит из абиссальных равнин, но эти плоские, пустынные регионы покрыты морскими отложениями и обычно придонная жизнь (бентос) здесь отсутствует. Глубоководные донные рыбы более распространены в каньонах или на скалах посреди равнин, где концентрируются сообщества беспозвоночных организмов. Подводные горы омываются глубоководными течениями, это становится причиной возникновения апвеллинга, который поддерживает жизнь донных рыб. Горные хребты могут делить подводные регионы на различные экосистемы[9].
В глубинах океана идёт непрерывный «морской снег», самопроизвольное осаждение детрита из верхних слоёв водной толщи. Он является производным жизнедеятельности организмов продуктивной эвфотической зоны. Морской снег включает мёртвый или умирающий планктон, простейших (диатомовые водоросли), песок, и прочую неорганическую пыль. По пути «снежинки» растут и за нескольких недель, пока не опустятся на дно океана, могут достигать нескольких сантиметров в диаметре. Тем не менее, большую часть органических компонентов морского снега потребляют микробы, зоопланктон и прочие животные-фильтраторы на протяжении первых 1000 метров их путешествия, то есть, в эпипелагиали. Таким образом, морской снег можно считать основой глубоководной мезопелагической и донной экосистем: поскольку солнечный свет не может проникнуть сквозь толщу воды, глубоководные организмы в качестве источника энергии используют морской снег.
Некоторые группы организмов, например, представителей семейств миктофовых, меламфаевых, фотихтиевых и топориковых, иногда называют псевдоокеаническими, поскольку они, обитая в открытом море, держатся вокруг структурных оазисов, подводных вершин или над материковым склоном. Подобные структуры также привлекают многочисленных хищников.
Характеристики
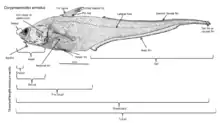
Глубоководные рыбы — одни из самых странных и неуловимых существ на Земле. В глубине обитает много необычных и неизученных животных. Они живут в полной темноте, поэтому, избегая опасности, и в поисках пищи и партнёра для размножения, не могут полагаться только на зрение. На больших глубинах преобладает свет синего спектра. Поэтому у глубоководных рыб диапазон воспринимаемого спектра сужен до 410—650 нм. У некоторых видов глаза имеют гигантские размеры и составляют 30—50 % длины головы (миктофовые, нансении, топорики-поллипнусы), а у других редуцированы или вообще отсутствуют (идиакантовые, ипноповые)[10]. Помимо зрения рыбы ориентируются на запах, электрорецепцию и изменение давления. Чувствительность к свету глаз некоторых видов в 100 раз превосходит человеческую[11].
По мере увеличения глубины давление увеличивается на 1 атмосферу каждые 10 м[12], тогда как концентрация пищи, содержание кислорода и интенсивность циркуляции воды снижаются[8]. У адаптированных к огромному давлению глубоководных рыб скелет и мускулатура развиты слабо. За счёт проницаемости тканей внутри тела рыбы давление равно давлению внешней среды. Поэтому при быстром подъёме на поверхность их тело раздувается, внутренности вылезают изо рта, а глаза выходят из орбит[13]. Проницаемость клеточных мембран повышает эффективность биологических функций, среди которых наиболее важна выработка белков; адаптацией организма к условиям окружающей среды является также повышение доли ненасыщенных жирных кислот в липидах клеточной мембраны[14]. У глубоководных рыб отличный от пелагических баланс метаболических реакций. Биохимические реакции сопровождаются изменением объёма. Если реакция приводит к увеличению объёма, она будет тормозиться давлением, а если к уменьшению, то она будет усилена[14]. Это означает, что метаболические реакции в той или иной степени должны уменьшить объём организма[15].
Более 50 % глубоководных рыб наряду с некоторыми видами креветок и кальмаров обладают биолюмнесценцией. Около 80 % из этих организмов имеют фотофоры, клетки, которые содержат бактерии, за счёт углеводов и кислорода из крови рыб вырабатывающие свет. Некоторые фотофоры имеют линзы, подобные линзам, содержащимся в глазах человека, которые регулируют интенсивность свечения. Рыбы затрачивают на испускание света всего 1 % энергии организма, тогда как оно выполняет несколько функций: с помощью света они ищут корм и привлекают добычу, подобно удильщикам; обозначают территорию в ходе патрулирования; общаются и находят партнёра для спаривания, а также отвлекают и временно ослепляют хищников. В мезопелагиали, куда проникает небольшое количество солнечного света, фотофоры на брюхе некоторых рыб маскируют их на фоне поверхности воды, делая невидимыми для хищников, плавающих ниже[11].

У некоторых глубоководных рыб часть жизненного цикла протекает на мелководье: там появляется на свет молодь, которая уходит на глубину по мере взросления. Вне зависимости от того, где находятся икра и личинки, всё это типично пелагические виды. Подобный планктонный, дрейфующий образ жизни требует нейтральной плавучести, поэтому в икре и плазме личинок присутствуют жировые капельки[16]. У взрослых особей имеются иные адаптации для сохранения положения в водной толще. В целом, вода выталкивает, поэтому организмы всплывают. Чтобы противодействовать выталкивающей силе, их плотность должна быть больше, чем у окружающей среды. Большая часть животной ткани плотнее воды, поэтому необходим уравновешивающий баланс[17]. Гидростатическую функцию у многих рыб выполняет плавательный пузырь, но у многих глубоководных рыб он отсутствует, а у большинства обладающих пузырем он не соединяется с кишечником при помощи протока[13]. У глубоководных рыб связывание и резервирование кислорода внутри плавательного пузыря, возможно, осуществляется липидами[15]. Например, у гоностомовых пузырь наполнен жиром[10]. Не имея плавательного пузыря, рыбы адаптировались к окружающей среде. Известно, что чем глубже среда обитания, тем более желеобразное тело у рыб и меньше доля костной структуры. Кроме того, плотность тела снижается за счёт повышенного содержания жира, и уменьшения веса скелета (меньший размер, толщина, содержание минеральных веществ и повышенная аккумуляция воды)[18]. Подобные характеристики делают обитателей глубин медлительными и менее подвижными по сравнению с пелагическими рыбами, живущими у поверхности воды.
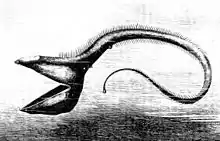
Отсутствие солнечного света на глубине делает фотосинтез невозможным, поэтому источником энергии глубоководным рыбам служит органика, опускающаяся сверху и, реже, гидротермальные источники срединно-океанических хребтов. Глубоководная зона менее богата питательными веществами, по сравнению с более мелкими слоями. Длинные чувствительные усики на нижней челюсти, например, как у долгохвостовых и трески, помогают искать пищу. Первые лучи спинных плавников удильщиков превратились в иллиций со светящейся приманкой. Огромная пасть, шарнирно сочленённые челюсти и острые зубы, как у мешкоротообразных, позволяют поймать и проглотить целиком крупную добычу[11].
Рыбы различных глубоководных пелагических и донных зон заметно отличаются друг от друга по поведению и строению. Группы сосуществующих видов в пределах каждой зоны функционируют аналогичным образом, например, небольшие мезопелагические фильтраторы, совершающие вертикальные миграции, батипелагические удильщики и глубоководные донные долгохвостовые[9].
Среди видов, обитающих на глубине, редко встречаются колючепёрые. Вероятно, глубоководные рыбы — достаточно древние и настолько хорошо приспособлены к окружающей среде, что проникновение современных рыб не увенчалось успехом[19]. Несколько глубоководных представителей колючепёрых принадлежат к древним отрядам бериксообразных и опахообразных. Большинство пелагических рыб, встречающихся на глубине, принадлежат к собственным отрядам, что предполагает длительную эволюцию в подобных условиях. И, напротив, глубоководные донные виды принадлежат к отрядам, которые включают многих рыб, обитающих на мелководье[20].
| Виды по зонам пелагиали | |
|---|---|
| Многие виды совершают суточные вертикальные миграции между зонами.
В этой таблице они перечислены по средней или более глубокой зоне основного пребывания | |
| Зона | Виды и группы видов: |
| Эпипелагическая[21] |
|
| Мезопелагческая | светящиеся анчоусы, опахи, алепизавры, опистопроктовые, малакосты, меламфаевые, саблезубые |
| Батипелагическая | В основном гоностомовые и удильщики. А также длиннорогие саблезубы, хаулиоды, живоглоты, гигантуровые, омосудовые, кинжалозубы, веретенниковые, угольные рыбы-сабли, циемовые, лофотовые, мешкоротообразные, китовидковые, топориковые |
| Бентопелагическая[21] | Особенно многочисленны долгохвостовые и бититовые |
| Бентическая | Камбалы, зеленоглазковые, бельдюги, угри, скаты, пинагоры и лопаточные нетопыри[21] |
| Сравнительное строение пелагических рыб | |||
|---|---|---|---|
| Параметр | Мезопелагические виды | Батипелагические виды | Глубоководные и бентические виды |
| мускулы | тело мускулистое | плохо развиты, тело дряблое | |
| скелет | крепкий | слабо окостеневший | |
| чешуя | да | нет | |
| нервная система | хорошо развита | только боковая линия и обоняние | |
| глаза | крупные и чувствительные | маленькие, иногда не функциональные | различные (от хорошо развитых до атрофированных) |
| фотофоры | часто имеются | часто имеются | обычно нет |
| жабры | хорошо развиты | ||
| почки | крупные | маленькие | |
| сердце | крупное | маленькое | |
| плавательный пузырь | имеется у рыб, совершающих вертикальные миграции | редуцирован или отсутствует | различный (от хорошо развитого до полного отсутствия) |
| размер | обычно менее 25 см | различный, виды длиной 1 м встречаются редко | |
Мезопелагические рыбы
Донные и придонные рыбы
Глубоководных донных рыб называют батидемерсальными. Они обитают за краем прибрежных бентических зон, в основном на материковом склоне и у континентального подножья, которое переходит в абиссальную равнину, встречаются у подводных вершин и островов. У этих рыб плотное тело и отрицательная плавучесть. Всю жизнь они проводят на дне. Одни виды охотятся из засады и способны зарываться в грунт, другие активно патрулируют дно в поисках пищи[22].
Примером рыб, способных зарываться в грунт, служат камбаловые и скаты. Камбалообразные — отряд лучепёрых рыб, которые ведут донных образ жизни, лежат и плавают на боку. Плавательный пузырь у них отсутствует. Глаза смещены на одну стороны тела. Личинки камбал первоначально плавают в толще воды, по мере развития их тело трансформируется, адаптируясь к жизни на дне[23]. У одних видов оба глаза расположены на левой стороне тела (арноглоссы), а у других — на правой (палтусы).
Ипноповые, живущие также на дне, выглядят совершенно иначе. Обычно они неподвижно стоят, опираясь на длинные утолщённые краевые лучи брюшных плавников и хвоста (иногда их называют рыбами-треногами). Они охотятся из засады на планктонных рачков. Длинные лучи грудных плавников функционируют подобно чувствительным антеннам, поскольку глаза у этих рыб развиты плохо[24].


Бентопелагические рыбы

Бентопелагические или придонные рыбы обитают в непосредственной близости у дна, питаются бентосом и бентопелагическим зоопланктоном[26]. Большинство демерсальных рыб относятся к бентопелагическим[27]. Их можно разделить на виды с крепким телом и дряблотелых. Дряблотелые бентопелагические виды схожи с батипелагическими, у них небольшая масса тела и низкий уровень метаболизма. Они затрачивают минимум энергии и охотятся из засады[28]. Примером такого типа служит Acanthonus armatus[29], хищник с крупной головой и телом, которое на 90 % состоит из воды. У этих рыб самые крупные глаза (отолиты) и самый маленький мозг относительно тела среди позвоночных[30].

Твёрдотелые бентопелагические рыбы — активные пловцы, которые энергично ищут добычу на дне. Иногда они живут вокруг подводных вершин с сильным течением[30]. Примером такого типа являются патагонский клыкач и атлантический большеголов. Ранее эти рыбы водились в изобилии и являлись ценным объектом промысла, их добывали ради вкусного плотного мяса[31][32].
У костистых бентопелагических рыб имеется плавательный пузырь. Типичные представители, ошибневые и долгохвостовые, довольно массивны — до 20 кг (чёрный конгрио); их длина достигает 2 м (малоглазый макрурус). Среди донно-придонных много трескообразных рыб, в частности моровых, спиношипообразных и галозавров[24][33].
Бентопелагические акулы, подобные глубоководным катранообразным, достигают нейтральной плавучести за счёт печени, богатой жиром[34]. Акулы хорошо приспособлены к довольно высокому давлению на глубине. Они попадаются на материковом склоне на глубине до 2000 м, где питаются падалью, в частности останками мёртвых китов. Однако для постоянного движения и сохранения жировых запасов им необходимо много энергии, которой недостаточно в олиготрофных условиях глубоководья[34].
Глубоководные скаты ведут бентопелагический образ жизни, у них, как у акул, крупная печень, поддерживающая их на плаву[34].
Глубоководные бентические рыбы
Глубоководные донные рыбы обитают за границей континентального шельфа. По сравнению с прибрежными видами они более разнообразны, поскольку в их среде обитания присутствуют различные условия. Бентические рыбы чаще попадаются и более разнообразны на континентальном склоне, где варьируется среда обитания и больше корма.
Типичные представители глубоководных донных рыб — ошибневые, долгохвостовые, угри, бельдюговые, зеленоглазковые, нетопырёвые и пинагоровые[3].
Самый глубоководный из известных ныне видов — Abyssobrotula galatheae, внешне похожие на угрей и совершенно слепые донные рыбы, которые питаются беспозвоночными[35].
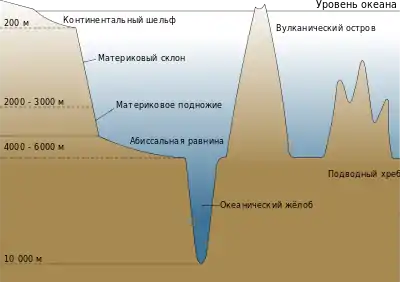
На больших глубинах дефицит пищи и чрезвычайно высокое давление ограничивает выживаемость рыб. Самая глубокая точка океана находится на глубине около 11 000 метров. Батипелагические рыбы обычно не встречаются ниже 3000 метров. Наибольшая глубина обитания донных рыб составляет 8370 м[35]. Возможно, экстремальное давление подавляет важнейшие функции ферментов[36].
У глубоководных бентических рыб, как правило, мускулистое тело и хорошо развитые органы. По строению они ближе к мезопелагическим, чем к батипелагическим рыбам, но они более разнообразны. У них обычно нет фотофор, у некоторых видов глаза и плавательный пузырь развиты, а у других отсутствуют. Размер также различен, однако длина редко превышает 1 м. Тело зачастую вытянутое и узкое, угревидное. Вероятно, это связано с удлинённой боковой линией, улавливающей низкочастотные звуки, с помощью которых некоторые рыбы привлекают половых партнёров[19]. Судя по той скорости, с которой глубоководные донные рыбы обнаруживают приманку, обоняние также играет важную роль в ориентации, наряду с осязанием и боковой линией[22].
Основу рациона глубоководных бентических рыб составляют беспозвоночные и падаль.
Как и в прибрежной зоне, донные рыбы глубоководья делятся на бентических с отрицательной и бентопелагических с нейтральной плавучестью тела[22].
По мере увеличения глубины падает количество доступной пищи. На глубине 1000 м биомасса планктона составляет 1 % от биомассы у поверхности воды, а на глубине 5000 м лишь 0,01 %[33]. Поскольку солнечный свет уже не проникает сквозь толщу воды, единственный источник энергии — органические вещества. Они попадают в глубинные зоны тремя способами.
Во-первых, органические вещества перемещаются с континентальной части суши через потоки речной воды, которые затем попадают в море и спускаются вдоль континентального шельфа и материкового склона. Во-вторых, в глубинах океана идёт непрерывный «морской снег», самопроизвольное осаждение детрита из верхних слоёв водной толщи. Он является производным жизнедеятельности организмов продуктивной эвфотической зоны. Морской снег включает мёртвый или умирающий планктон, простейших (диатомовые водоросли), фекалии, песок, сажу и прочую неорганическую пыль. Третий источник энергии обеспечивают совершающие вертикальные миграции мезопелагические рыбы. Особенностью этих механизмов является то, что количество питательных веществ, которые попадают к донным рыбам и беспозвоночным, постепенно уменьшается по мере удаления от континентальных береговых линий[2].
Несмотря на скудость кормовой базы, среди глубоководных донных рыб существует определённая пищевая специализация. Например, они отличаются по размеру рта, который определяет размер возможной добычи. Некоторые виды питаются бентопелагическими организмами. Другие поедают животных, обитающих на дне (эпифауна), или зарывающихся в грунт (инфауна). У последних в желудках наблюдается большое количество грунта. Инфауна служит вторичным источником пищи для падальщиков, подобных синафобранховым[37].

Некоторые виды питаются падалью. Видеозаписи показывают, что когда мёртвая рыба опускается на дно, к ней устремляются позвоночные и беспозвоночные падальщики. Если тело крупное, они погружаются в него целиком и выедают изнутри. В свою очередь они привлекают хищников, подобных долгохвостовым, которые начинают охотиться на питающихся падальщиков. Существует также пищевая специализация на основе вертикального распределения. Наиболее многочисленные в верхней части материкового склона виды рыб, такие как слитножаберные угри и нитепёрые налимы[38], питаются в основном эпипелагическими рыбами. Но в целом рацион большинства глубоководных демерсальных видов состоит из беспозвоночных[37][39].
Примечания
- Ильмаст Н. В. Введение в ихтиологию. — Петрозаводск: Карельский научный центр РАН, 2005. — ISBN 5-9274-0196-1.
- Moyle, P. B. and Cech, J. J., 2004, pp. 594.
- Moyle, P. B. and Cech, J. J., 2004, pp. 587.
- Trujillo, Alan P., and Harold V. Thurman, 2011, pp. 354.
- Trujillo, Alan P., and Harold V. Thurman, 2011, pp. 365.
- Trujillo, Alan P., and Harold V. Thurman, 2011, pp. 457, 460.
- P. J. Cook, Chris Carleton. Continental Shelf Limits: The Scientific and Legal Interface. — 2000. — ISBN 0-19-511782-4.
- Moyle, P. B. and Cech, J. J., 2004, pp. 585.
- Moyle, P. B. and Cech, J. J., 2004, pp. 591.
- А. А. Иванов. Физиология рыб / Под ред. С. Н. Шестах. — М.: Мир, 2003. — 284 с. — (Учебники и учебные пособия для студентов высших учебных заведений). — 5000 экз. — ISBN 5-03-003564-8.
- Trujillo, Alan P., and Harold V. Thurman, 2011, pp. 414—415.
- Wharton, David, 2002, pp. 198.
- Никольский Г. В. Биология рыб. — Советская наука, 1944. — С. 9.
- Wharton, David, 2002, pp. 199, 201—202.
- Завалеева С., Сизова Е., Чиркова Е. Эволюционно-функциональная морфология животных. — Science, 2016.
- Randall, David J., and Anthony Peter Farrell, 1997, pp. 217.
- Randall, David J., and Anthony Peter Farrell, 1997, pp. 195.
- Randall, David J., and Anthony Peter Farrell, 1997, pp. 196, 225.
- R. L. Haedrich. Deep-water fishes: evolution and adaptation in the earth's largest living spaces* (англ.) // Journal of Fish Biology. — 1996-12-01. — Vol. 49. — P. 40–53. — ISSN 1095-8649. — doi:10.1111/j.1095-8649.1996.tb06066.x.
- Moyle, P. B. and Cech, J. J., 2004, pp. 586.
- Moyle, P. B. and Cech, J. J., 2004, pp. 571.
- Moyle, P. B. and Cech, J. J., 2004, pp. 588.
- E. A. Fairchild, W. H. Howell. Factors affecting the post-release survival of cultured juvenile Pseudopleuronectes americanus (англ.) // Journal of Fish Biology. — 2004-12-01. — Vol. 65. — P. 69–87. — ISSN 1095-8649. — doi:10.1111/j.0022-1112.2004.00529.x.
- Н. В. Парин. Рыбы открытого океана / Под редакцией чл.- корреспондента АН СССР А. П. Андрияшева. — М.: Наука, 1988. — ISBN 5-02-005246-9.
- Bathypterois grallator (англ.) в базе данных FishBase.
- J. Mauchline, J. D. M. Gordon. Foraging strategies of deep-sea fish // Marine Ecology Progress Series. — № 27. — P. 227—238. — doi:10.3354/meps027227.
- Walrond C Carl. Coastal fish - Fish of the open sea floor. Te Ara - the Encyclopedia of New Zealand.
- J. A. Koslow. Energetic and life-history patterns of deep-sea benthic, benthopelagic and seamount-associated fish (англ.) // Journal of Fish Biology. — 1996-12-01. — Vol. 49. — P. 54–74. — ISSN 1095-8649. — doi:10.1111/j.1095-8649.1996.tb06067.x.
- Acanthonus armatus (англ.) в базе данных FishBase.
- M. L. Fine, M. H. Horn, B. Cox. Acanthonus armatus, a Deep-Sea Teleost Fish with a Minute Brain and Large Ears (англ.) // Proceedings of the Royal Society of London B: Biological Sciences. — 1987-03-23. — Vol. 230, iss. 1259. — P. 257—265. — ISSN 0962-8452. — doi:10.1098/rspb.1987.0018.
- Промыслы клыкача. Регулятивная система. Commission for the Conservation of Antarctic Marine Living Resources.
- Атлантический большеголов. www.inokean.ru. Дата обращения: 7 февраля 2016.
- Bone Q. and Moore R. H., 2008, pp. 43.
- Bone Q. and Moore R. H., 2008, pp. 42.
- Nielsen J. G. The deepest living fish Abyssobrotula galatheae: a new genus and species of oviparous ophidioids (Pisces, Brotulidae) // Galathea Report. — 1977. — Vol. 14. — P. 41—48.
- Ryan P. Deep-sea creatures (англ.). Te Ara Encyclopedia of New Zealand. New Zealand Ministry for Culture and Heritage Te Manatu Taonga. Дата обращения: 5 февраля 2016.
- G. R. Sedberry, J. A. Musick. Feeding strategies of some demersal fishes of the continental slope and rise off the Mid-Atlantic Coast of the USA (англ.) // Marine Biology. — 1978-12-01. — Vol. 44, iss. 4. — P. 357–375. — ISSN 0025-3162. — doi:10.1007/BF00390900.
- Phycis chesteri (англ.) в базе данных FishBase.
- Moyle, P. B. and Cech, J. J., 2004, pp. 595.
Литература
- Ильмаст Н. В. Введение в ихтиологию. — Петрозаводск: Карельский научный центр РАН, 2005. — ISBN 5-9274-0196-1.
- Trujillo, Alan P., and Harold V. Thurman. Essentials of Oceanography 10th ed.. — Boston: Prentice Hall, 2011. — ISBN 978-0321668127.
- Moyle, P. B. and Cech, J. J. Fishes, An Introduction to Ichthyology (5th ed.). — Benjamin Cummings, 2004. — ISBN 978-0-13-100847-2.
- Wharton, David. Life at the Limits: Organisms in Extreme Environments. — Cambridge, UK: Cambridge UP, 2002. — P. 198. — ISBN 978-0521782128.
- Randall, David J., and Anthony Peter Farrell. Deep-sea Fishes. — San Diego: Academic, 1997. — ISBN 978-0123504401.
- Bone Q. and Moore R. H. Biology of Fishes. — Taylor & Francis Group, 2008. — ISBN 978-0-415-37562-7.
| В библиографических каталогах |
|---|