Велоцирапторины
Велоцираптори́ны[1] (лат. Velociraptorinae) — подсемейство тероподовых динозавров в составе семейства дромеозаврид.
| † Велоцирапторины | ||||||
|---|---|---|---|---|---|---|
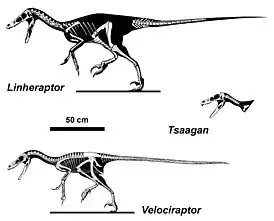 Скелеты велоцирапторин | ||||||
| Научная классификация | ||||||
|
Домен: Царство: Подцарство: Без ранга: Без ранга: Тип: Подтип: Инфратип: Надкласс: Клада: Клада: Клада: Клада: Клада: Надотряд: Отряд: Подотряд: Клада: Клада: Инфраотряд: Семейство: Подсемейство: † Велоцирапторины |
||||||
| Международное научное название | ||||||
| Velociraptorinae Barsbold, 1983 | ||||||
| Типовой род | ||||||
| ||||||
Описание
Самыми ранними представителями, вероятно, являлись Nuthetes, найденный в Великобритании, и дейноних из Северной Америки. Однако, было обнаружено несколько неопределённых велоцирапторин, чьи остатки датируются киммериджской эпохой (верхний юрский период). Эти окаменелости были обнаружены в карьере Лангерберг, Окер, недалеко от города Гослар, Германия[2].
В 2007 году палеонтологи изучили кости передних конечностей велоцираптора и обнаружили небольшие бугорки на поверхности, известные как маховые бугорки. Эта же особенность присутствует на некоторых птичьих костях и представляет собой точку крепления для мощных вторичных перьев крыла. Это открытие явилось первым прямым свидетельством того, что велоцирапторины, как и все прочие манирапторы, имели перья[3].
В то время как большинство велоцирапторин были маленькими животными, по крайней мере один вид мог достигать гигантских размеров, сопоставимых с теми, которые были найдены среди дромеозаврин. До сих пор этот гигантский безымянный велоцирапторин известен только по отдельным зубам, найденным на острове Уайт, Англия. Это зубы животного размером с ютараптора, но они, по-видимому, принадлежали велоцирапторину, судя по форме зубов и форме их кромок[4].
Отличительные черты анатомии
Диагноз — это описание анатомических особенностей организма (или группы), которые в совокупности отличают его от всех других организмов. Диагноз таксономической группы организмов включает в себя признаки, известные как синапоморфии, которые разделяют два или более организма в группе и, как полагают, присутствуют у их последнего общего предка.
Согласно Карри, велоцирапторин можно отличить по следующим характеристикам[5]:
- дромеозавриды с верхне- и нижнечелюстными зубами, имеющими пилообразные насечки на передних краях, которые значительно меньше на задних зубах, и которые имеют второй премаксилярный зуб, который значительно больше третьего и четвёртого премаксилярных зубов;
- дромеозавриды, имеющие сплющенные носовые отверстия, которые выглядят таковыми при осмотре сбоку.
Согласно Тёрнеру, представителей подсемейства Velociraptorinae можно отличить по следующим характеристикам[6]:
- заднее отверстие базисфеноидной ямки разделено на два маленьких круглых отверстия около тонкой удлинённой кости;
- дорсальная барабанная ямка представляет собой глубокую, постерио-латерально направленную полость;
- наличие полостей (плевроцелей) во всех спинных позвонках.
Классификация

Будучи созданным в 1983 году Р. Барсболдом, подсемейство Velociraptorinae задумывалось как группа, включающая велоцираптора и его ближайших родственников. Так было до 1998 года, когда Пол Серено определил её как кладу, содержащую всех дромеозаврид, которые ближе к дромеозавру, чем к велоцираптору. Ряд более поздних исследований обозначил Velociraptorinae как группу дромеозаврид, тесно связанных с велоцираптором, состав которой широко варьировался в зависимости от того, какие виды считать велоцирапторинами и какие из них более базальны или ближе к дромеозавру.
Новас и Пол в 2005 году создали отдельную кладу велоцирапторин, близкую к типовому виду, куда вошли велоцираптор, дейноних и таксон, впоследствии названный Tsaagan. Проведённый Тёрнером в 2012 году кладистический анализ подтвердил версию о монофилии велоцирапторин[6]. Однако, некоторые исследователи (Longrich & Currie, 2009) в составе велоцирапторин создали особую группу дромеозаврид, где дейноних был признан не-велоцирапториновым, не-дромеозавриновым эудромеозавром, а Saurornitholestes стал членом более базальной группы, названной Saurornitholestinae. Более тщательный анализ 2013 года сделал некоторых традиционных велоцирапторин, таких как Tsaagan, более базальными, чем велоцираптор, в то время как другие были более тесно связаны с дромеозавром, что делало их дромеозавринами. Это же исследование показало, что Balaur, ранее считавшийся велоцирапторином, оказался принадлежащим группе Avialae[7].
Присутствие остатков этого животного в верхнемеловых отложениях Северной Америки, наряду с Acheroraptor и Dakotaraptor, указывает на то, что велоцирапторины продолжали эволюционировать и диверсифицироваться после своего кампан-маастрихтского переселения из Азии. Поскольку эти таксоны не образуют монофилетическую кладу, предполагается, что в кампанскую и маастрихтскую эпохи присутствовало несколько родов дромеозаврид, в том числе как минимум 2 рода в северных, и 1 — в южных пределах Ларамидии. Эти линии следовали определёнными эволюционными путями, по-видимому, заполняя похожие экологические ниши в соответствующих экосистемах. Филогенетический анализ, выполненный при описании велоцирапторина Dineobellator, предполагает, по крайней мере, 4 кампанские линии, и, по крайней мере, 2—3 из них остаются в Северной Америке в маастрихте[8].
Филогенетический анализ А. Аверьянова и А. Лопатина (2021) восстановил следующую кладограмму велоцирапторин при описании дромеозаврида кансайгната[9][10]:
| Dromaeosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Примечания
- Курочкин Е. Н., Лопатин А. В. Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Часть 2 / Лопатин А. В.. — М.: Геос, 2012. — Т. 2. — С. 207. — 419 (154 рис., 26 фототабл.) с. — ISBN 978-5-89118-594-4.
- van der Lubbe T., Richter U., Knotschke N. Velociraptorine dromaeosaurid teeth from the Kimmeridgian (Late Jurassic) of Germany (англ.) // Acta Palaeontologica Polonica. — 2009. — Vol. 54, no. 3. — P. 401—408.
- Turner A. H., Makovicky P. J., Norell M. A. Feather quill knobs in the dinosaur Velociraptor (англ.) // Science. — 2007. — Vol. 317, no. 5745. — P. 1721. — doi:10.1126/science.1145076.
- Naish D. H., Martill, D. M. Saurischian dinosaurs: theropods // Dinosaurs of the Isle of Wight. — The Palaeontological Association, Field Guides to Fossils, 2001. — P. 10, 242—309.
- Currie P. J. New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda) (англ.) // Journal of Vertebrate Paleontology. — 1995. — No. 15. — P. 576—591. — doi:10.1080/02724634.1995.10011250.
- Turner A. H., Makovicky P. J., Norell M. A. A Review of Dromaeosaurid Systematics and Paravian Phylogeny (англ.) // Bulletin of the American Museum of Natural History. — 2012. — Vol. 371, no. 1. — doi:10.1206/748.1.
- Godefroit P., Cau A., Hu D.-Y., Escuillié F., Wu W., Dyke G. A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds (англ.) // Nature. — 2013. — Vol. 498, no. 7454. — P. 359—362. — doi:10.1038/nature12168.
- Jasinski S. E., Sullivan R. M., Dodson P. New dromaeosaurid dinosaur (Theropoda, Dromaeosauridae) from New Mexico and biodiversity of dromaeosaurids at the end of the Cretaceous (англ.) // Scientific Reports. — 2020. — Vol. 10, no. 5105. — doi:10.1038/s41598-020-61480-7.
- Новый позднемеловой хищный динозавр из Средней Азии (англ.). Палеонтологический институт им. А. А. Борисяка РАН. Дата обращения: 26 июня 2021. Архивировано 26 июня 2021 года.
- Аверьянов А. О., Лопатин А. В. Новый хищный динозавр (Theropoda, Dromaeosauridae) из позднего мела Таджикистана // Доклады Российской академии наук. Науки о Земле. — 2021. — Т. 499, № 1. — С. 49—53. — ISSN 2686-7397. — doi:10.31857/S2686739721070045.