R-тела
R-тела — это нерастворимые «ленты» из белков, синтезируемые некоторыми видами бактерий. Обычно в цитоплазме бактерий эти ленты плотно свернуты в цилиндрические структуры[1]. Изначально они были обнаружены в каппа-частицах — бактериальных эндосимбионах инфузорий рода Paramecium. Они или кодирующие их гены встречаются и у некоторых видов свободноживущих бактерий, функции R-тел у этих бактерий неизвестны. R-тела можно разделить на пять различных групп, отличающихся по размеру, морфологии лент и механизму раскручивания R-тел[2].
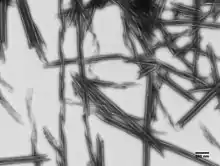
Морфология, сборка и удлинение
При нейтральном pH R-тела типа 51 напоминают свёрнутую ленту 500 нм в диаметре и примерно 400 нм в высоту[1]. Кодируется единственным опероном из четырёх открытых рамок считывания[3][4]. R-тела состоят из двух маленьких структурных белков, RebA и RebB[5]. Третий белок, RebC, необходим для ковалентного соединения двух структурных белков в высокомолекулярную структуру, которая выглядит как лестница на белковом форезе[5].
При низком pH R-тела разворачиваются начиная от центра, образуя пустую трубку с заострёнными концами длиной до 20 микрометров[6].
Функции
R-тела обеспечивают убийство чувствительных штаммов инфузорий другими инфузориями. Когда каппа-частицы от инфузории-«убийцы» попадают в организм инфузории-«жертвы», R-тела под действием кислой среды в её пищеварительной вакуоли разворачиваются, образуя трубочку диаметром 165 нм и длиной 20 мкм (для R-тел типа 51). При этом R-тела разрывают мембраны каппа-частицы и пищеварительной вакуоли, смешивая цитоплазмы бактерии (каппа-частицы) и инфузории-жертвы[7]. Последующая смерть инфузории предположительно вызвана попаданием в её цитоплазму токсина, содержащегося в каппа-частицах. Инфузория не умирает, когда поглощает очищенные R-тела или кишечных палочек, экспрессирующих белки R-тел. Таким образом, R-тела не убивают клетку, а скорее играют роль средства доставки других молекул[3][8]. R-тела могут быть использованы в качестве настраиваемых устройств для приложения в клеточной инженерии, так как R-тела типа 51 являются натуральными биологическими машинами, переходящими от скрученной формы к развернутой в ответ на изменения pH. Они могут возвращаться к своей свернутой форме при повышении pH. Также они могут работать в жестких условиях, таких как высокая температура, присутствие солей и детергентов. Процесс развертывания R-тел представляет собой простое и действенное решение задачи разрыва мембраны; таким образом, он является многообещающим клиническим инструментом для высвобождения и взаимопроникновения содержимого разных мембранных отсеков друг в друга[9].
Примечания
- F R Pond, I Gibson, J Lalucat,R L Quackenbush. R-body-producing bacteria. (англ.) // Microbiology and Molecular Biology Reviews : journal. — American Society for Microbiology, 1989. — 1 March (vol. 53, no. 1). — P. 25—67. — ISSN 0146-0749. — PMID 2651865.
- Raymann, Kasie; Bobay, Louis-Marie; Doak, Thomas G.; Lynch, Michael; Gribaldo, Simonetta. A genomic survey of Reb homologs suggests widespread occurrence of R-bodies in proteobacteria (англ.) // G3: Genes, Genomes, Genetics : journal. — 2013. — 1 March (vol. 3, no. 3). — P. 505—516. — ISSN 2160-1836. — doi:10.1534/g3.112.005231. — PMID 23450193.
- Kanabrocki, J. A.; Quackenbush, R. L.; Pond, F. R. Organization and expression of genetic determinants for synthesis and assembly of type 51 R bodies (англ.) // Journal of Bacteriology : journal. — 1986. — 1 October (vol. 168, no. 1). — P. 40—48. — ISSN 0021-9193. — PMID 3759909.
- Jeblick, Jörn; Kusch, Jürgen. Sequence, transcription activity, and evolutionary origin of the R-body coding plasmid pKAP298 from the intracellular parasitic bacterium Caedibacter taeniospiralis (англ.) // Journal of Molecular Evolution : journal. — 2005. — 1 February (vol. 60, no. 2). — P. 164—173. — ISSN 0022-2844. — doi:10.1007/s00239-004-0002-2. — PMID 15785846.
- Heruth, D. P.; Pond, F. R.; Dilts, J. A.; Quackenbush, R. L. Characterization of genetic determinants for R body synthesis and assembly in Caedibacter taeniospiralis 47 and 116 (англ.) // Journal of Bacteriology : journal. — 1994. — 1 June (vol. 176, no. 12). — P. 3559—3567. — ISSN 0021-9193. — PMID 8206833.
- Preer, John R.; Hufnagel, Linda A.; Preer, Louise B. Structure and behavior of R bodies from killer paramecia (англ.) // Journal of Ultrastructure Research : journal. — 1966. — 1 April (vol. 15, no. 1). — P. 131—143. — doi:10.1016/S0022-5320(66)80100-4. — PMID 5936490.
- Mueller, Jo Anne. Vitally stained kappa in Paramecium aurelia (англ.) // Journal of Experimental Zoology : journal. — 1965. — 1 December (vol. 160, no. 3). — P. 369—372. — ISSN 1097-010X. — doi:10.1002/jez.1401600314. — PMID 4160786.
- Schrallhammer, Martina; Galati, Stefano; Altenbuchner, Josef; Schweikert, Michael; Görtz, Hans-Dieter; Petroni, Giulio. Tracing the role of R-bodies in the killer trait: absence of toxicity of R-body producing recombinant E. coli on paramecia (англ.) // European Journal of Protistology : journal. — 2012. — 1 November (vol. 48, no. 4). — P. 290—296. — ISSN 1618-0429. — doi:10.1016/j.ejop.2012.01.008. — PMID 22356923.
- Jessica K. Polka, Pamela A. Silver. A. A Tunable Protein Piston That Breaks Membranes to Release Encapsulated Cargo (англ.) // ACS Synthetic Biology : journal. — 2016. — 15 April (vol. 5, no. 4). — P. 303—311. — doi:10.1021/acssynbio.5b00237. — PMID 26814170.