Генная конверсия
Генная конверсия — замена одной последовательности ДНК на гомологичную последовательность, так что последовательности становятся идентичными. Конверсия гена может быть либо аллельной, что означает, что один аллель гена заменяет другой аллель того же гена, либо неаллельной/эктопической[1], при которой одна паралогичная ДНК-последовательность преобразует другую.[2]
Паралогичными называют такие последовательности, которые присутствуют в одном и том же геноме, а возникают в эволюции путём дупликации (удвоения) первичной последовательности. Паралогичные гены образуют семейства генов, и события конверсии между генами одного семейства достаточно часты[1]. Конверсия может происходить между последовательностями функциональных генов, которые кодируют белки; также между псевдогенами — участками генома, которые гомологичны по последовательности какому-то функциональному гену, но утратили способность обеспечивать наработку продукта в виде РНК или белка; может также происходить конверсия функционального гена на последовательность псевдогена (и это в некоторых случаях приводит к возникновению патологии развития эмбриона из-за утраты работоспособности необходимого гена), либо наоборот, конверсия псевдогена по образцу последовательности функционального гена.
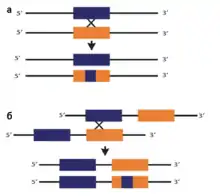
Конверсия может затрагивать участки разной протяженности, иногда конверсией затрагивается одна часть гена при сохранении уникальности последовательностей в другой части (в этом случае говорят о сегментной конверсии гена).
С точки зрения поддержания стабильности генома конверсия играет двоякую роль: с одной стороны, она может помочь заместить мутантную версию гена с патогенными свойствами на нормальную версию того же гена. Но с другой стороны, может происходить и противоположная конверсия с замещением нормальной версии на патогенную. В этом случае мы можем получить гомозиготизацию генотипа по патогенному рецессивному варианту и проявление рецессивого заболевания у ребёнка, вопреки ожиданиям на основании генотипирования родителей. Таким образом, генная конверсия может нарушать стандартные менделевские расщепления при наследовании. Конверсия и гомозиготизация по мутациям в онкогенах отмечалась и в клетках опухолей.
Механизм
В основе генной конверсии лежит процесс репарации молекулы ДНК по механизму гомологичной рекомбинации. Этот механизм задействован в процессе восстановления нити ДНК, в которой произошел двуцепочечный разрыв. Когда в ходе гомологичной рекомбинации происходит комплементарное спаривание между двумя гомологичными последовательностями, в местах несоответствия двух цепочек образуется гетеродуплекс, что запускает активацию репарации неспаренных оснований. Эта система и корректирует одну цепочку по матрице другой как по образцу.
Здесь можно отметить, что вероятность замен одних нуклеотидов на другие неодинакова. Если в одной из цепочек находится нуклеотид Г или Ц, а во второй А или Т, то вероятность замещения нуклеотида во второй цепочке по матрице первой выше, чем наоборот. Это явление называют асимметрией конверсии (по-английски conversion bias). Это приводит к тому, что, например, в области горячих точек рекомбинации, где события конверсии происходят особенно часто, наблюдается повышенная концентрация нуклеотидов Г и Ц по сравнению с геномом в целом[3]. Также на вероятность конверсии той или иной цепочки может влиять то, какой из участков более активно вовлекается в процесс транскрипции: выше вероятность коррекции менее активной цепочки по матрице более активной, чем наоборот[4]. Адаптивный смысл такой работы системы репарации достаточно очевиден: слабее экспрессирующийся фрагмент ДНК, вероятно, может содержать мутацию, нарушающую его работоспособность, а конверсия замещает этот вариант на более работоспособный.
Регулярная конверсия генов
В ряде случаев организмам требуется иметь очень высокое разнообразие определенных классов последовательностей. В первую очередь это касается работы систем, задействованных в различении «свой-чужой». К примеру, нам необходимо строить разнообразные иммуноглобулины и антитела, обеспечивающие защиту от разнообразных вторгающихся инфекционных микроорганизмов, сами микроорганизмы же, со своей стороны, нуждаются в возможности быстро заменять поверхностные антигены, дабы успешно избегать атаки иммунной системы хозяина. В подобных системах генная конверсия активно используется клетками в качестве механизма генерирования огромного разнообразия рабочих последовательностей. Суть механизма следующая. Необходимый организму белок производится за счёт работы определенного гена, который находится под активным промотором и экспрессируется. Также в геноме присутствуют множественные и разнообразные паралогичные последовательности, которые не экспрессируются, но периодически в случайном порядке используются в качестве матрицы для конверсии активной последовательности. Именно так, к примеру, генерируется разнообразие сменяющих друг друга поверхностных антигенов паразита трипаносомы (одноклеточного возбудителя сонной болезни)[5]. В этом случае весь процесс запрограммирован на генетическом уровне. Цепочка событий начинается с образования двуцепочечного разрыва вблизи активного гена, но этот разрыв появляется не случайно как повреждение, а вносится высокоспецифичным ферментом эндонуклеазой, который активируется в ответ на определенные внутриклеточные сигналы.
Примечания
- Frequent nonallelic gene conversion on the human lineage and its effect on the divergence of gene duplicates (англ.) // Proceedings of the National Academy of Sciences. — 2017-11-28. — Vol. 114, iss. 48. — P. 12779–12784. — ISSN 1091-6490 0027-8424, 1091-6490. — doi:10.1073/pnas.1708151114.
- Jian-Min Chen, David N. Cooper, Nadia Chuzhanova, Claude Férec, George P. Patrinos. Gene conversion: mechanisms, evolution and human disease (англ.) // Nature Reviews Genetics. — 2007-09-11. — Vol. 8, iss. 10. — P. 762–775. — ISSN 1471-0064 1471-0056, 1471-0064. — doi:10.1038/nrg2193.
- Laurent Duret, Nicolas Galtier. Biased Gene Conversion and the Evolution of Mammalian Genomic Landscapes (англ.) // Annual Review of Genomics and Human Genetics. — 2009-09. — Vol. 10, iss. 1. — P. 285–311. — ISSN 1545-293X 1527-8204, 1545-293X. — doi:10.1146/annurev-genom-082908-150001.
- Ezra Schildkraut, Cheryl A. Miller, Jac A. Nickoloff. Transcription of a Donor Enhances Its Use during Double-Strand Break-Induced Gene Conversion in Human Cells (англ.) // Molecular and Cellular Biology. — 2006-4. — Vol. 26, iss. 8. — P. 3098–3105. — ISSN 0270-7306. — doi:10.1128/MCB.26.8.3098-3105.2006.
- David Horn. Antigenic variation in African trypanosomes (англ.) // Molecular and Biochemical Parasitology. — 2014-7. — Vol. 195, iss. 2. — P. 123–129. — ISSN 0166-6851. — doi:10.1016/j.molbiopara.2014.05.001.