Воротная теория боли
Воротная теория боли (теория воротного контроля боли) (англ. gate control theory) — теория, объясняющая механизм регуляции болевой чувствительности с помощью механизма ворот в задних рогах спинного мозга. Теория была предложена Р. Мелзаком и П. Уоллом в 1965 году.
История
Р. Мелзак и П. Уолл при построении своей теории опирались на уже существующие теорию специфичности и теорию паттернов[1]. Теория специфичности гласит, что существуют особые болевые рецепторы и волокна болевой чувствительности, которые передают болевые импульсы в центр боли в головном мозге без изменений. А болевая чувствительность — это отдельная модальность аналогично зрительной и слуховой модальностям[1].
Существует группа теорий, которые можно объединить в теорию паттернов[1]. Часть теорий основываются на предположении о том, что качественно различные кожные ощущения (ощущения сдавливания, температурная чувствительность, болевая чувствительность формируются благодаря разным пространственно-временным паттернам нервных импульсов[2]. Другая часть теорий постулируют в качестве основного механизма болевой чувствительности механизм суммации нервных импульсов[1], аналогичный тем, которые описаны Д. Хеббом[3] и Р. Джерардом[4]. С теориями суммации также связана теория, постулирующая существование миелинизированных (толстых) и немиелинизированных (тонких) проводящих волокон. При нормальном их функционировании толстые волокна оказывают тормозящее действие на тонкие. В случае гипералгезии этого не происходит и тонкие проводящие волокна доминируют над толстыми[5].
Таким образом, толстым и тонким проводящим волокнам не приписывается специфическая чувствительность, но они специфичны по функции — быстрые волокна способны изменять сенсорный вход для импульсов, передающихся тонкими волокнами[1][5].
Воротная теория
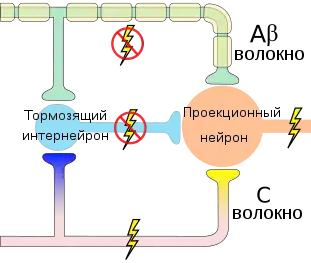
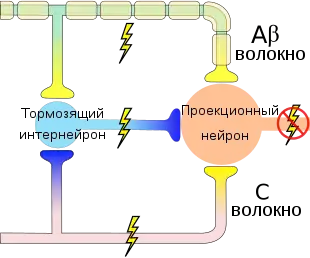
Согласно воротной теории боли, болевая чувствительность детерминирована взаимодействием трёх систем: системы воротного контроля, системы триггера центрального контроля и системы действия.
Система воротного контроля
Желатинозная субстанция спинного мозга работает как ворота, контролирующие сенсорный вход от периферических нервных волокон. Она регулирует возбуждающее действие толстых и тонких волокон на вставочные клетки спинного мозга[7].
Импульсы, приходящие по толстым волокнам максимально эффективно действуют в начале, а затем их работа тормозится желатинозной субстанцией, поскольку они оказывают возбуждающее действие на клетки желатинозной субстанции, а желатинозная субстанция, в свою очередь, тормозит передачу импульса на терминалях и толстых, и тонких волокон. Импульсы же, передающиеся по тонким волокнам, со временем усиливаются желатинозной субстанцией, поскольку они оказывают на клетки желатинозной субстанции тормозящее действие[7].
Тонкие немиелинизированные волокна способны к тоническому напряжению и медленно адаптируются. Поток импульсов в спинной мозг по ним не прекращается. Они держат ворота в относительно открытом состоянии. Толстые миелинизированные волокна большую часть времени находятся в неактивном состоянии и при отсутствии изменения стимуляции адаптируются[1].
При предъявлении стимула повышается число активных волокон. Сенсорные входы от тонких и толстых ворот уравновешивают друг друга, и выход вставочных клеток растет медленно. Затем, если уровень стимуляции остается неизменным, пропорция активных толстых и тонких волокон смещается — толстые волокна инактивируются, соответственно, ворота открываются, и выход вставочных клеток увеличивается быстрее[1].
Таким образом, на выход вставочных клеток влияют общее число активированных волокон и соотношение активности толстых и тонких волокон. Выход вставочных клеток может значительно отличаться от суммы афферентных импульсов, поступающих на них[1].
Система триггера центрального контроля
Нисходящие влияния головного мозга способны влиять на проводимость афферентных нервных волокон. В случае болевой чувствительности этот центральный контроль опосредствован воротной системой контроля[8].
По быстрым волокнам большого диаметра импульсы идут не только в желатинозную субстанцию, но и в головной мозг, где происходит избирательная активация мозговых процессов, способных влиять на работу воротного контроля, а также на информацию, которая продолжает поступать по более медленным волокнам[1].
Система действия
Когда величина сенсорного входа на вставочные клетки превышает критический уровень, запускается система действия. Таким образом, после повреждения кожи происходит следующая последовательность действий: испуг, рефлекс сгибания, постуральная корректировка, вокализация, изменение положения головы и глаз с целью оценки поврежденного участка, вегетативный ответ, актуализация предшествующего опыта и предсказание последствий такой стимуляции и другие паттерны поведения[1].
Применение теории
Воротная теория позволяет объяснить феномен гипералгезии. При травматических повреждениях или некоторых невропатиях большая часть толстых волокон разрушается. Соответственно, ворота оказываются открыты, что создает благоприятные условия для пространственной суммации импульсов. В таких условиях даже легкое касание может вызывать болевые ощущения[1]. Таким же образом можно объяснить феномен спонтанной боли. Тонкие волокна проявляют спонтанную активность, следовательно, открывают ворота. Благодаря суммации эти спонтанные импульсы вызывают боль[1].
Задержки ощущений после стимуляции могут возникать при поражениях, когда общее число периферических нервов уменьшилось. Вставочным клеткам нужно время, чтобы перестроиться, и сначала общего количества поступающих импульсов не хватает для превышения критического уровня[1].
Феномен отражённой боли тоже можно объяснить с точки зрения воротной теории. Каждая вставочная клетка имеет ограниченное рецептивное поле, но на нее могут влиять и импульсы, идущие от более широких участков тела[9]. В норме воротный механизм не пропускает импульсы, способные вызвать болевые ощущения на этих обширных участках. При потере волокон ворота оказываются открыты[1].
Таким же образом теория объясняет и феномен триггерных точек[1].
Облегчение боли
Боль можно уменьшить или уменьшением входа от тонких волокон, или увеличением входа от толстых волокон[1]. Например, боль при каузалгии можно уменьшить, если поместить конечность в теплую ванну и массировать ее[10]. Приятные прикосновения увеличивают вход от толстых волокон, соответственно, ворота закрываются.
Значение воротной теории
Несмотря на многие ошибочные моменты, воротная теория значительно продвинула изучение боли в целом[11]. Если более ранние теории считали, что ощущение боли напрямую зависит от интенсивности поступающих сигналов, то Р. Мелзак и П. Уолл впервые заявили, что избирательные и интегративные действия предшествуют первой синаптической передаче импульса[12].
Схема, предложенная Р. Мелзаком и П. Уоллом, представляет собой первое приближение основных механизмов, работающих, в спинном мозге. Она может быть легко понятна лицам, имеющим косвенное отношение к изучению непосредственно боли[13].
Критика
Теория не учитывала известные факты о стимульной специфичности периферических нервных волокон. Более приближено к реальности представление о двух группах нервных волокон: одна группа получает количественную информацию о стимулах и учитывает паттерн, а другая — специфичные для отдельных видов стимуляции волокна[12].
Воротная теория боли не рассматривает факт разных болевых ощущений при раздражении разных тканей, тогда как боль зависит не только от типа волокон, но и от их расположения в тканях[12]. Можно выделить три слоя внутри кожи: внешний слой вызывает зуд и жгучую боль, следующий слой вызывает поверхностную яркую боль, а дерма — ноющую боль[14].
Также существуют исследования, опровергающие наличие двух противоположных эффектов при активации толстых и тонких волокон[12].
Дальнейшее развитие теории
Большинство предположений теории было позднее подтверждено, хотя они и нуждаются в дальнейших уточнениях[13]. Часть критических замечаний были учтены П. Уоллом в более поздней статье[15]. В 1990 году Р. Мелзак расширил теорию до теории нейроматрикса[16].
См. также
Примечания
- R. Melzack, P. D. Wall. Pain Mechanisms: A New Theory (англ.) // Science. — 1965-11-19. — Vol. 150, iss. 3699. — P. 971–978. — ISSN 1095-9203 0036-8075, 1095-9203. — doi:10.1126/science.150.3699.971.
- John Paul Nafe. The Pressure, Pain, and Temperature Senses. (англ.) // A handbook of general experimental psychology. / Carl Murchison. — Worcester: Clark University Press, 1934. — P. 1037–1087. — doi:10.1037/11374-019.
- 1. Hebb D.O. The Organization of Behavior, Wiley: New York. — 1949.
- 1. Gerard R. The Phenomenon of Causalgia // Anesthesiology. — 1951. — January (Vol. 12). — pp. 1 — 10.
- Noordenbos W. Problems pertaining to the transmission of nerve impulses which give rise to pain: Preliminary statement // Elsevier Science Publishers, Amsterdam. — 1959.
- Kandel, Eric R. Principles of Neural Science / Eric R. Kandel, James H. Schwartz, Thomas M. Jessell. — 4th. — New York : McGraw-Hill, 2000. — P. 482–486. — ISBN 0-8385-7701-6.
- L. M. Mendell, P. D. Wall. Presynaptic hyperpolarization: a role for fine afferent fibres (англ.) // The Journal of Physiology. — 1964-07-01. — Vol. 172, iss. 2. — P. 274–294. — doi:10.1113/jphysiol.1964.sp007417.
- 1. Hagbarth K. E., Kerr D. I. B. Central influences on spinal afferent conduction // Neurophysoil. — 1954. — (Vol. 17) — pp. 295—307.
- Mendell L. M., Wall P. D. Responses of single dorsal cord cells to peripheral cutaneous unmyelinated fibres // Nature. — 1965. — (Vol. 206, no. 4979) — pp. 97 — 99.
- W. K. Livingston. The vicious circle in causalgia (англ.) // Annals of the New York Academy of Sciences. — 1948-10. — Vol. 50, iss. 4 Teleological. — P. 247–258. — ISSN 1749-6632 0077-8923, 1749-6632. — doi:10.1111/j.1749-6632.1948.tb39855.x.
- Lorne M. Mendell. Constructing and deconstructing the gate theory of pain: (англ.) // Pain. — 2014-02. — Vol. 155, iss. 2. — P. 210–216. — ISSN 0304-3959. — doi:10.1016/j.pain.2013.12.010.
- P. W. Nathan. The gate-control theory of pain: a critical review (англ.) // Brain. — 1976. — Vol. 99, iss. 1. — P. 123–158. — ISSN 1460-2156 0006-8950, 1460-2156. — doi:10.1093/brain/99.1.123.
- Sufka K. J., Price D. D. Gate control theory reconsidered // Brain and Mind. — 2002. — (Vol.3) — 277—290.
- Keele C. A., Armstrong D. Substances Producing Pain and Itch // London: Edward Arnold. — 1964.
- P.D. Wall. The gate control theory of pain mechanisms: a re-examination and re-statement (англ.) // Brain. — 1978. — Vol. 101, iss. 1. — P. 1–18. — ISSN 1460-2156 0006-8950, 1460-2156. — doi:10.1093/brain/101.1.1.
- Ronald Melzack. Phantom limbs and the concept of a neuromatrix (англ.) // Trends in Neurosciences. — 1990-03. — Vol. 13, iss. 3. — P. 88–92. — doi:10.1016/0166-2236(90)90179-E.